Larvae of pandalid shrimp
물렁가시붉은새우(
따라서 본 연구에서는 물렁가시붉은새우 조에아 유생의 저서 생활을 하는 생태적 특징을 고려하여 가진 영양강화한 알테미아, 바닥으로 공급되는 이매패류의 육질부 그리고 새우용 배합사료를 이용하여 사육 초기 적정 먹이를 규명하기 위한 사육 실험을 실시하여 보고하고자 한다.
2013년 2월 2-3일, 이틀 동안 부화한 물렁가시붉은새우(
1개월 간의 사육에서 얻어진 성장한 물렁가시붉은새우 치하는 개체수를 측정하여 생존율을 계산한 후, 임의의 10마리씩을 전장 및 미각장 측정을 위해 일부의 사육수와 함께 얼음물에 담궜다가 0.01 mm까지 디지털 버니어켈리퍼스(Mytutoyo, Japan)로 측정하였다. 성장 측정을 마친 치하는 개체별로 구분하여 60℃ 열풍건조기(Daesan, Korea)에서 24시간 방치 후 전자저울(Precisa, Swiss)로 0.1 mg까지 측정하였다.
실험에 사용된 새우의 사료는 영양강화한 알테미아, 다진 바지락 살 그리고 치하용 사료를 사용하였다, 바지락은 NFRDI(2009)의 수산물 성분표(2009)를 참고하여 일반성분과 지방산 및 아미노산을 mg/g dry matter로 환산하여 확인하였으며, 치하용 사료와 영양강화한 알테미아는 동결건조 후 2 mg 이하로 정량된 시료를 자동아미노산분석기(L-8800, Hitachi, Japan)로 분석하였으며 Folch et al. (1957)의 방법으로 지질을 추출하여 Parrish (1987)의 방법에 따라 유리지방산을 methylation 시켜서 auto-sampler가 장착된 Gas chromatography (6890N, Agilent, U.S.A.)로 분리분석(Kwon et al., 2013a, 2013b)하였다. 실험 종료 시 치하의 핵산분석을 위한 시료는 각 실험구의 각 반복수조에서 임의로 2마리씩 채취하여 분석 시까지 -25℃에 보관하였다. 채취된 치하는 micro grinders (Kontes PELLET PESTLE®)를 이용하여 1,800 rpm에서 1분간 균질화하여 핵산분석용 시료로 사용하였다. RNA와 DNA는 3 M KOH 와 5% perchloric acid (PCA)를 용매로 하여 추출하였다. 그리고 추출된 RNA와 DNA 수용액은 흡광계(Thermo, U.S.A.)로 260 nm의 파장값을 Peragón et al. (2001)과 Fukuda et al. (1986)에 방법에 따라 정량하였다. 치하의 수용성 단백질은 각 시료의 균질액 중 일정량을 원심분리한 후 상등액을 Bradford (1976)의 Bio-red protein assay 법으로 측정하여 albumin으로 정량하였다.
실험을 통해 얻어진 1개월 사육 치하의 생존율, 전장, 미각장, 건조중량, 단백질, 핵산비 및 RNA와 DNA 함량은 SPSS program (Ver 19.0)을 이용하여 ANOVA test를 실시한 후 Duncan의 다중검정을 통해 평균간의 유의성을 95% 범위에서 분석하였다(Duncan, 1955).
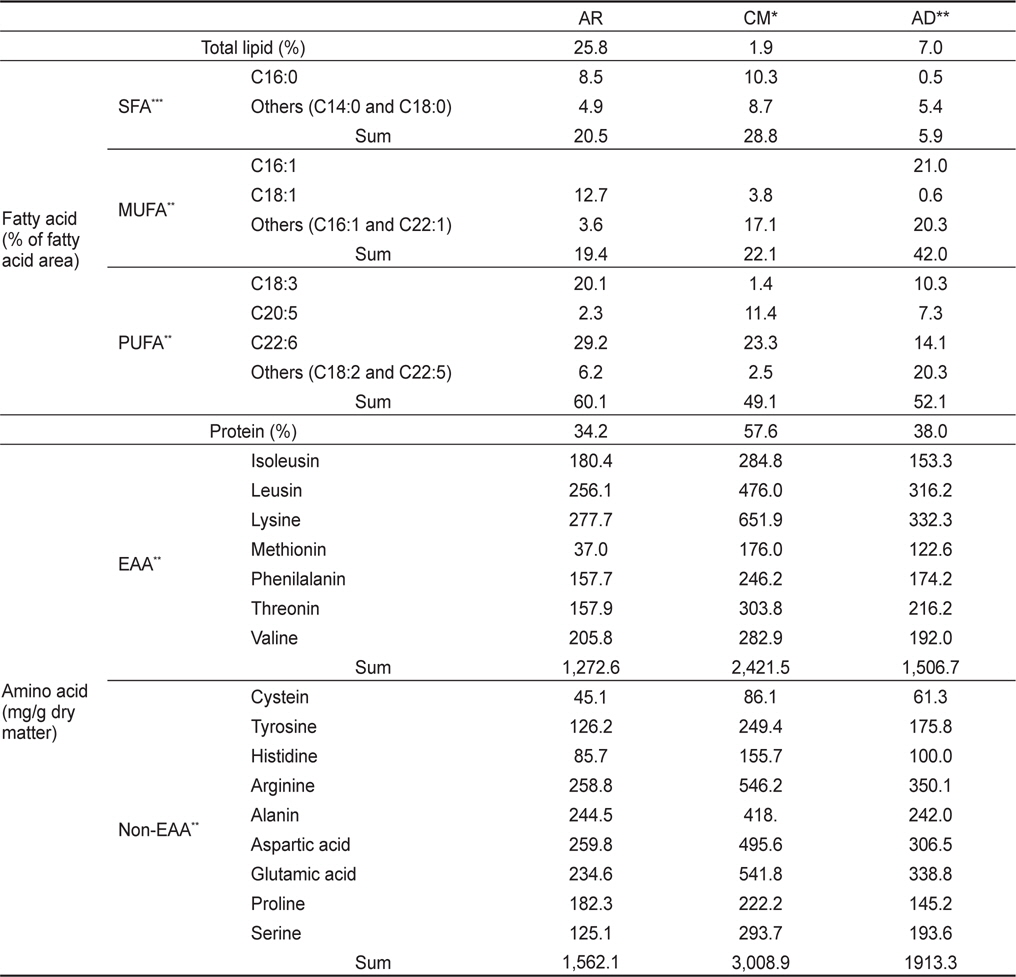
물렁가시붉은새우 유생의 성장 및 생존율 실험에 사용된 먹이의 총 지질과 지방산 조성은 Table 1에 나타내었다. AR구는 총지질이 25.8%였으며, 이중 포화지방산이 20.5%, 단일불포화지방산이 19.4%였고 고도불포화지방산은 60.1%로 높았다. CM구는 총 지질함량 1.9% 중 28.8%가 포화지방산, 22.1%가 단일불포화지방산이었고, 49.1%가 고도불포화지방산이었다. AD구는 7.0%의 총 지질 함량이었다. 지방산 조성으로는 영양강화한 알테미아가 EPA 함량은 2.32%, DHA 함량이 29.2%를 차지하였다. 조갯살인 경우 EPA가 9.6%였으며, DHA 함량이 23.3%로 조사되었다.
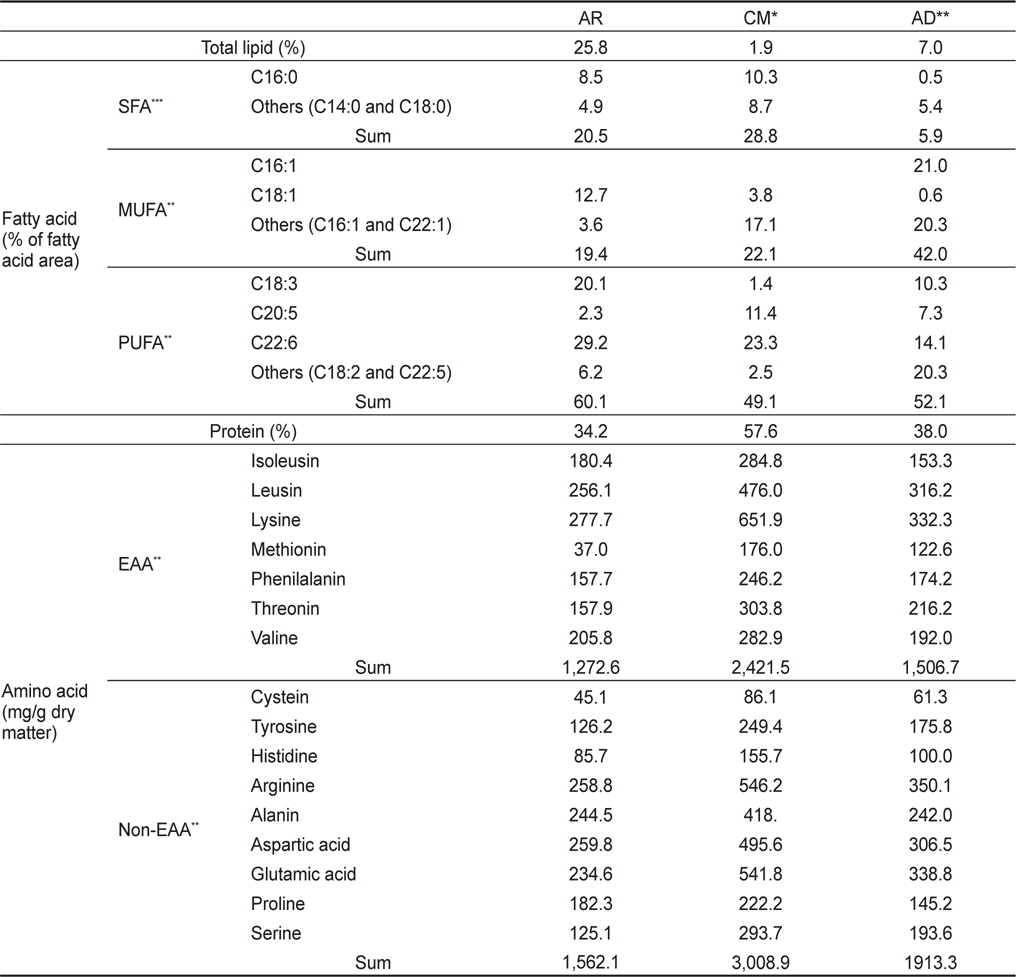
Fatty acid composition (% of total fatty acids) and protein-bound amino acid (mg/g dry matter) of enriched Artemia sp. for 24 hours (AR), clam Tapes philippinarum meat (CM) and artificial crumbled shrimp diets (AD) supplied for the Pandalid shrimp, Pandalopsis japonica larvae for a month
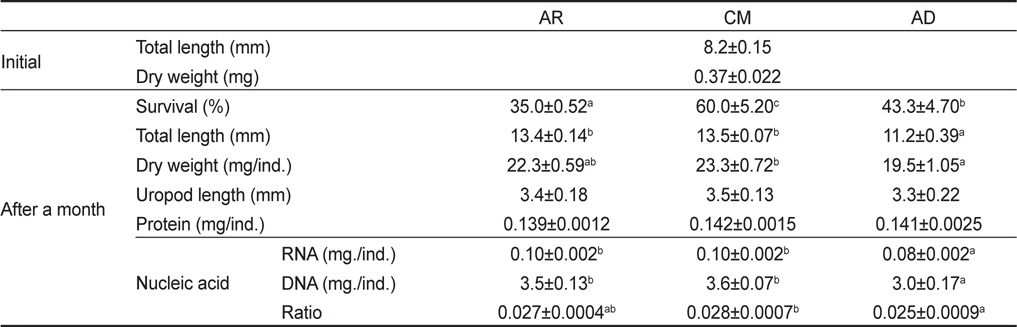
1개월간 성장한 물렁가시붉은새우 치하의 생존, 성장, 수용성단백질 및 핵산 함량은 Table 2에 나타내었다. 생존율은 CM에서 60.0±5.20%로 유의적으로 가장 높게 나타났으며(P<0.05), AR에서 가장 낮은 35.0±0.52%로 조사되었다. 전장은 13.5±0.07 mm로 CM에서 유의적으로 가장 높았지만(P<0.05), AR구와 유의적인 차이는 없었다(P>0.05). 건조중량과 미각장은 모든 실험구에서 19.5-23.3 mg과 3.3-3.5 mm로 차이를 나타내지 않았다(P>0.05). 개체별 단백질 함량 또한 0.139-0.142 mg/ind.로 실험구간 유의적인 차이를 보이지 않았다(P>0.05). 핵산에서 RNA 함량은 AR과 CM에서 각각 0.10±0.002 mg/ind.와 0.10±0.002 mg/ind.로 0.08±0.002 mg/ind.의 AD보다 유의적으로 높았다(P<0.05). DNA 함량은 AR과 CM에서 각각 3.5±0.13 mg/ind.와 3.7±0.07 mg/ind.로 AD의 3.0±0.17 mg/ind.보다 유의적으로 높았다(P<0.05). 또한 RNA/DNA ratio는 CM에서 0.028±0.0007로 유의적으로 가장 높았지만(P<0.05), AR의 0.027±0.0004와 유의적인 차이는 없었다(P>0.05).
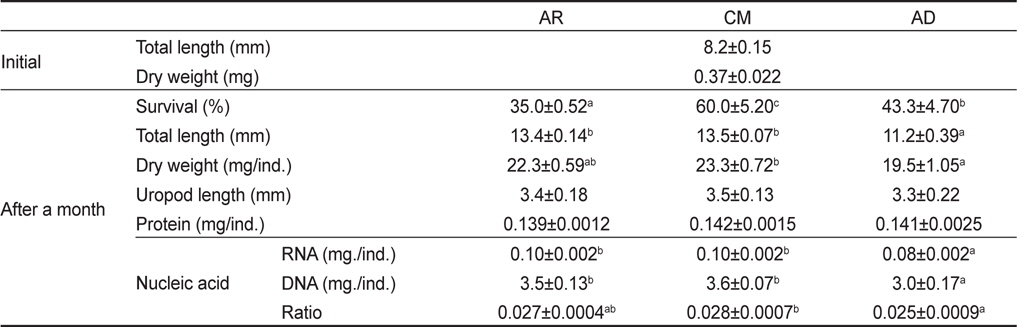
Total length (mm), uropod length (mm), wet weight (mg), survival (%), protein contents (mg/ind.), RNA contents (mg/ind.), DNA contents (mg/ind.) and RNA/DNA ratio of Pandalopsis japonica post-larva fed on different three diets (AR, CM and AD)1
동해안 유용새우류 중 물렁가시붉은새우(
사료는 주로 흰다리새우,
새우 유생의 식성이 실험실에서와 자연에서 다를 수는 있지만, Levay et al. (2001)은 장의 형태를 근거로 새우,
이것을 뒷받침할 근거로 핵산관련 지수를 사용할 수 있다. 한 개체에서의 RNA 함량은 세포 내 활성을 직접적으로 가리키는 것이기 때문에(Fukuda et al., 1986) 사료공급구에서 낮은 RNA 함량은 섭취 혹은 섭취 후 소화/흡수된 영양분의 양이 적다는 것을 의미한다. 그리고 사료 공급구에서 DNA 함량이 낮은 것은 낮은 세포활성으로 세포분열이 적었기 때문에 단위 세포 내에 일정량 존재하는 이 함량이 낮게 조사되었던 것이다. 결국 단위세포 당 세포의 활성도의 지표가 되는 RNA/DNA ratio가 사료 공급구에서 낮게 나타난 것은 낮은 세포 활성으로 인한 체성장이 늦다는 것을 나타내는 결과이다. 이 ratio와 체성장 경향은 일치한다.
결과적으로 본 연구에서 영양적으로 낮지 않은 사료와 알테미아 공급구에서 보인 결과는 의미가 상반된다고 할 수 있다 첫번째로 AR구의 성장은 양호했지만 생존율이 낮은 이유는 부유성 알테미아를 충분히 섭취하기에는 물렁가시붉은새우 초기 유생들의 활력이 낮다는 것을 의미한다. 두 번째로 사료 공급구의 낮은 성장과 생존율은 초기 유생들의 낮은 소화력으로 인해 흡수할 수 있는 영양분이 적어서 낮았다고 판단된다. 하지만 위(functional stomach)의 기능이 활발해지는 조에아 2기 혹은 메갈로파 단계에서는 미립자사료의 소화흡수력이 높아졌을 가능성이 있기 때문에 실험 초기 높은 폐사율이 있었을 가능성을 시사하는 부분이다. 결국 조개를 다져서 공급하는 것은 물렁가시 붉은새우들의 유생의 낮은 소화력과 부족한 포식능력을 고려하 였을 때 가장 적합한 먹이라고 판단된다. 하지만 방류 전까지 성장을 시키기에는 다소 먹이공급상의 불편한 부분이 있기 때문에 배합사료와 생먹이를 함께 공급하는 방법으로 개선해야 할 여지가 있다고 판단된다.