We examined seasonal variation in the structure of marine macroalgal communities at five sites around the Byeonsan Peninsula, Korea, from January to October 2011. Sixty eight species were identified, including 11 green, 17 brown and 40 red algae. Species richness was highest at Gyeokpo (51 species) followed by Sambal (47), Gosapo (34), Byeonsan (33), and Habgoo (18). Seaweed biomass ranged from 12.98 to 145.33 g dry wt/㎡, with maximum and minimum biomasses at Gyeokpo and Habgoo, respectively.
해조류는 선캄브리아기부터 존재하였던 가장 원시적인 식생그룹으로 해산식물종의 90%을 차지하며 해양먹이사슬에서 일차생산자로서 그리고 경제적으로 중요한 구성원이다(Manilal et al., 2010; Satheesh and Wesley, 2012; Silva et al., 2012). 해조류 군집은 어 · 패류를 포함한 수많은 해양생물의 은신처, 산란장 및 먹이장과 같은 생태적 서비스를 제공하므로, 연안생태계에서 먹이사슬의 상위에 있는 해양생물의 종다양성과 풍도, 나아가 어장형성에 영향을 준다(Wan et al., 2009; Janiak and Whitlatch, 2012; Satheesh and Wesley, 2012). 일차생산자인 해조류는 연안의 도시화와 산업화로 인해 해양에 유입 되는 증가된 무기 및 유기 오염물을 제거하는 생물여과자(biofilter)로서, 연안 환경을 모니터링하는 생물지표자(biological indicator)로 사용된다(Worm et al., 2000; Wells et al., 2007; Scherner et al., 2013).
이외에도 해조류는 식료품, 의약품, 해조 콜로이드산업, 화장품의 원료와 동물의 사료로서 다양한 산업과 문화 발전에 긍정적인 영향을 주는 중요한 수산식물이다(Zemke-White and Ohno, 1999; Koch et al., 2013). 하지만, 연안에 공업지대와 도시가 형성되면서 인간활동에 의해 오염과 부영양화가 진행되어 해조류의 종다양성과 생물량은 지속적으로 감소하고 이러한 환경에 잘 적응하는 갈파래류(
우리나라 서해안은 황해난류의 영향을 받으며, 리아스식 해안을 따라 하구, 만, 연성 및 경성 조간대 등 다양한 환경으로 구성되어있다(Jung et al., 2010). 또한, 갯펄로 구성된 연성 저질이 발달되고 조수 간만의 차가 커서 부유성 물질이 많아 탁도가 높으며 암반에는 많은 굴이 피복하고 남해·동해안에 비하여 해조류 생물량과 종다양도가 현저하게 낮은 것으로 알려져있다(Yang and Kim, 2009). 서해안 해조류는 Kang (1966)이 한국산 해조류의 지리적 분포 연구에서 140여 종으로 기록함을 시작으로, 해조상과 군집구조에 관한 생태 연구, 수평 · 수직 분포, 기능형군 분석, 생물량 및 다양도지수 등 다양한 생태학적 분석이 시도 되었다(Lee et al., 2007; Yoo et al., 2007; Choi et al., 2008).
서해안 변산반도는 국립공원지역으로 화려한 경관과 해수욕장이 잘 발달되어 다양한 해안 레저와 관광을 위해 방문객이 증가하면서 유·무기 오염물의 해양 투기량도 급증하고 있으며, 새만금 방조제의 완성으로 인하여 환경변화가 심한 지역이다(Kim et al., 2009; Choi et al., 2013). 따라서, 동해안과 남해안에 비해 해조류 서식환경이 열악한 서해안 변산반도의 환경변화와 기후변화에 따른 해조류 종조성과 군집구조의 변화 파악 및 건강한 연안생태계 유지를 위한 기초생산자의 보존 및 복원을 위한 기초자료 확보가 매우 중요한 시점이다.
따라서, 본 연구는 변산반도의 생태학적 특성과 생물자원인 해조류의 군집특성을 파악하기 위하여 변산반도 5개 정점에 대한 계절별 해조류의 종다양성, 생물량, 우점종 및 수직분포를 확인하기 위해 수행하였다.
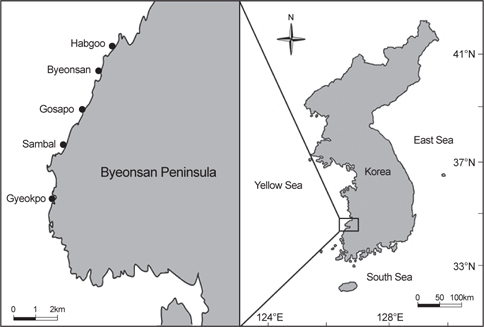
전라북도 변산반도의 5개 정점(합구, 변산, 고사포, 삼발, 격포)에서 2011년 1월부터 10월까지 계절별로 해조류를 정량 및 정성적으로 채집하였다(Fig. 1). 정량 채집은 조사지점의 해조 생육 상한선을 기준으로 저조선까지 가상의 line transect를 설치한 후 10×10 cm 으로 구획된 방형구(50×50 cm)를 조간대 상, 중, 하부에 각각 3개, 총 9개의 방형구를 놓고 Saito and Atobe (1970)의 방법에 의해 출현한 해조류의 빈도(F)와 피도(C)를 야장에 기록한 후 방형구내의 출현한 모든 해조류를 끌칼을 이용하여 전량 채집하였다. 해조상은 조사해역에 서식하는 모든 해조류를 정성 채집하여 확인했으며, 정량 및 정성 채집된 해조류는 현장에서 포르말린 – 해수용액 (5–10%)으로 고정시켜 실험실로 운반한 후 현미경을 사용하여 동정하였고, 출현종의 학명 및 국명은 Lee and Kang (2002)에 따랐다.
정량 채집된 해조류는 담수로 수회 세척하여 모래 및 불순물을 제거하고 동정한 후 60°C로 설정된 건조기에서 7일간 건조한 후 건중량을 측정하였고, 생물량(g/m2)으로 환산하였다. 피도는 현장에서 기록한 야장을 근거로 단위면적당 피복 백분율로써, 빈도는 전체 소방형구(25개)에 대한 각 종이 출현한 방형구수의 비로 계산하였으며 정점의 우점종을 판단하는 중요도(importance value)는 상대피도(RC)와 상대빈도(RF)의 산술 평균으로 나타냈다(Mueller-Dombois and Ellenberg, 1974). 해조류의 수평분포지수는 분류군별로 구분한 후 갈조류에 대한 녹조류의 비(C/P), 갈조류에 대한 홍조류의 비(R/P) 및 갈조류에 대한 홍조류와 녹조류 합의 비(R+C)/P로 계산하였다(Feldmann, 1937; Segawa, 1956; Cheney, 1977). 계절별로 출현한 해조류의 종별 평균 생물량과 종수를 근거로 하여 풍도지수(richness index, R), Shannon’s 다양도지수(diversity index, H’), 균등도지수(evenness index, J’), 우점도지수(dominance index)를 구하고, 각 정점의 유사도(similarity)를 분석하였다. 출현한 해조류의 우점도는 K-dominance 곡선으로 비교하였으며(Lambshead et al., 1983), 군집지수의 산출 및 도식화는 PRIMER version 6 (Clarke and Gorley, 2006)을 이용하였다.
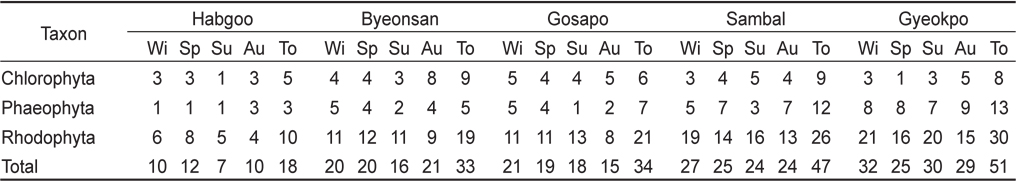
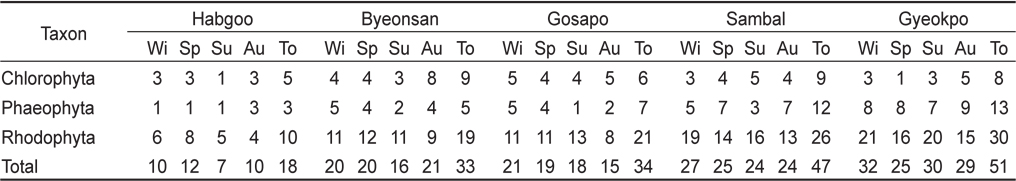
본 연구기간에 변산반도의 5개 정점(합구, 변산, 고사포, 삼발, 격포)에서 출현한 해조류는 총 68종으로 분류군별로 녹조류가 11종(16.18%), 갈조류가 17종(25.00%), 홍조류가 40종(58.82%)을 차지하여 홍조류의 출현종수가 최대였고, 갈조류와 녹조류의 순으로 나타났다(Table 1). 정점별로 일년간 출현한 해조류의 총수는 18–51종으로 격포에서 최대, 합구에서 최소였으며 변산반도 전체 조사 정점의 계절별 출현종수는 겨울 42종, 봄 43, 여름 40종 그리고 가을에 41종으로 봄에 최대였고 여름에 최소였다(Table 1).
The number of macroalgal species observed at the five study sites of Byeonsan Peninsula, Korea (Wi, Winter; Sp, Spring; Su, Summer; Au, Autumn; To, Total)
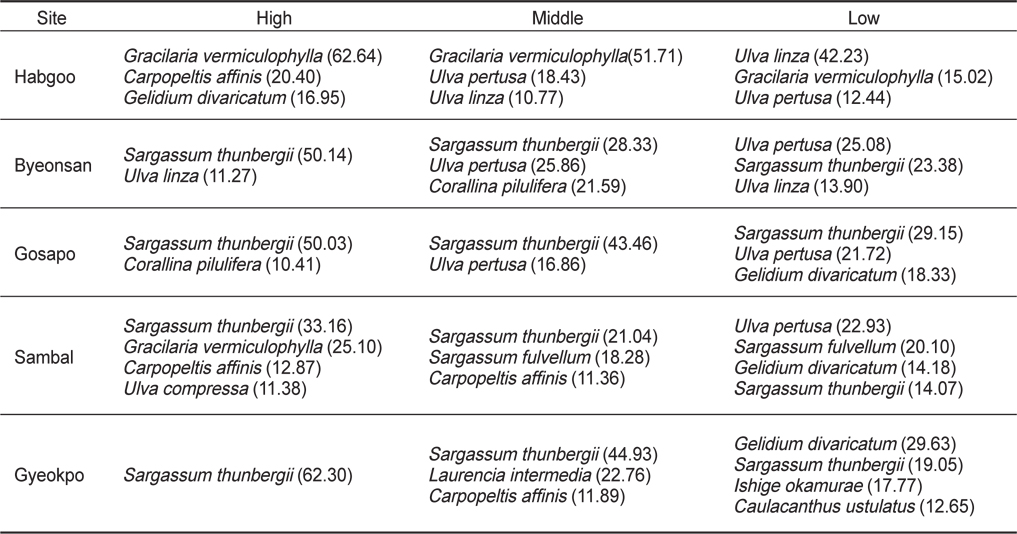
연구 정점별 해조상을 보면, 합구에서 출현한 해조류는 총 18종(녹조 5, 갈조 3, 홍조 10종)이었으며, 홍조류가 55.55%로 가장 높았다. 합구의 계절별 출현종은 7–12종으로 봄에 최대였고 여름에 최소였으며, 지충이(
변산반도 5개 정점에서 모두 출현한 종은 15종이었고, 분류군별로는 녹조류 4종(납작파래, 잎파래, 모란갈파래, 구멍갈파래), 갈조류 3종(세가닥갯쇠털, 참모자반, 지충이)과 홍조류 8종(방사무늬김, 애기우뭇가사리, 까막살, 애기가시덤불, 꼬시래기, 비단풀, 새빨간검둥이, 모로우붉은실)으로 확인되었다.
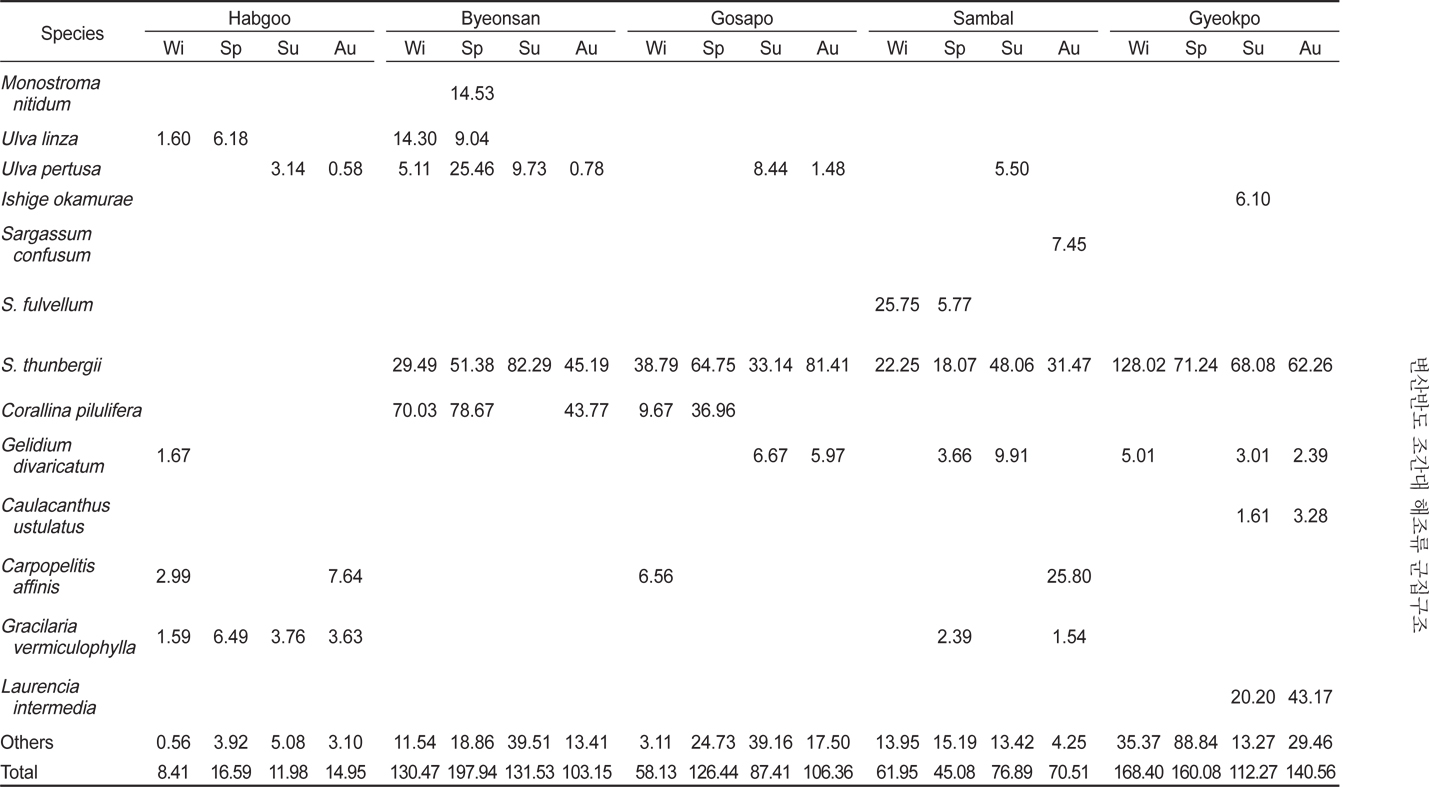
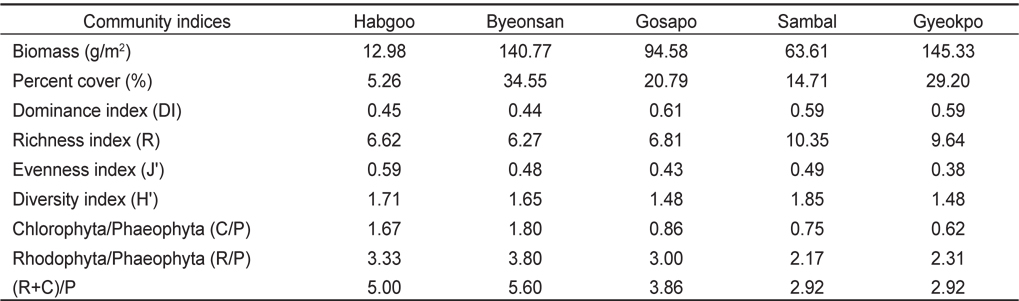
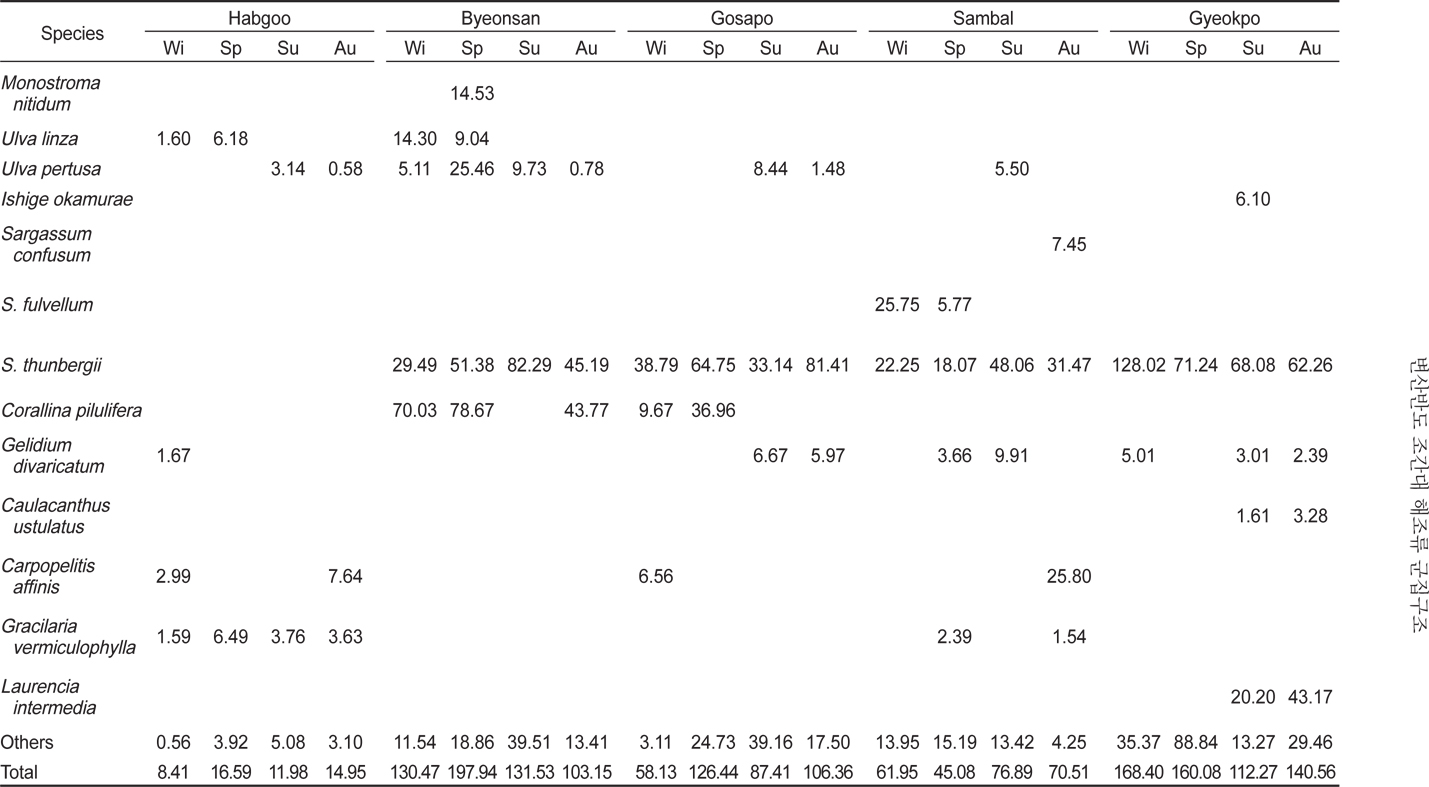
변산반도 5개 정점의 조간대에 서식하는 해조류의 연평균 생물량(g dry wt/㎡)은 91.45 g/㎡이고, 정점별 12.98-145.33 g/㎡로 합구에서 최소였고, 격포에서 최대로 나타났다(Table 2). 합구에서 해조류 생물량은 연평균 12.98 g/㎡(계절별 8.41- 16.95 g/㎡)였고, 조위별로는 조간대 상부에서 2.04 g/㎡, 중부에서 19.38 g/㎡, 하부에서 18.55 g/㎡로 중 · 하부에서 높았다. 변산에서 생물량은 연평균 140.77 g/㎡ (계절별 103.15- 197.94 g/㎡ ; 조위별 73.65-197.45 g/㎡)로서 상부에서 가장 낮았고 중부에서 최대였으며, 고사포에서는 연평균 94.58 g/㎡(계절별 58.13–126.44 g/㎡ ; 조위별 90.91-117.33 g/㎡)로서 중부에서 최대였다. 삼발에서 생물량은 94.58 g/㎡이었고 계절별로 45.08-76.89 g/㎡, 조위별 33.83-94.92 g/㎡로 상부에서 최대였다. 격포에서 생물량은 145.32 g/㎡로서 여름에 최소였고 겨울에 최대였으며(112.27-168.40 g/㎡), 조위별로는 79.22-211.69 g/㎡으로 하부에서 가장 낮고 중부에서 최대였다(Table 2).
Marine algal biomass (g dry wt/ ㎡ ) occurred at five study sites in Byeonsan Peninsula, Korea (Wi, Winter; Sp, Spring; Su, Summer; Au, Autumn)
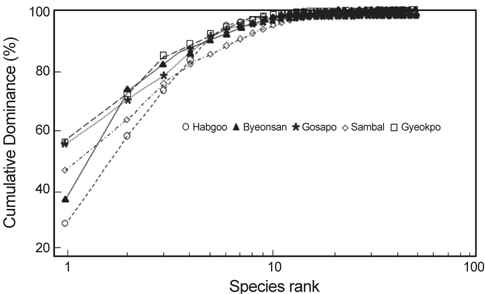
정점별 해조류의 연평균 생물량과 종다양성을 근거로 K– dominance곡선을 도식화하면, 합구에서는 까막살, 꼬시래기와 잎파래가 전체 생물량의 76.43%를 차지하였고 변산에서는 작은구슬산호말, 지충이와 구멍갈파래가 81.53%, 고사포에서는 우점종인 지충이, 작은구슬산호말과 까막살이 80.58%를 차지하였다. 또한, 삼발에서는 지충이, 까막살과 참모자반이 생물량의 76.42%를, 그리고 격포에서는 지충이, 작은구슬산호말과 검은서실이 84.62%를 보였다(Fig. 2).
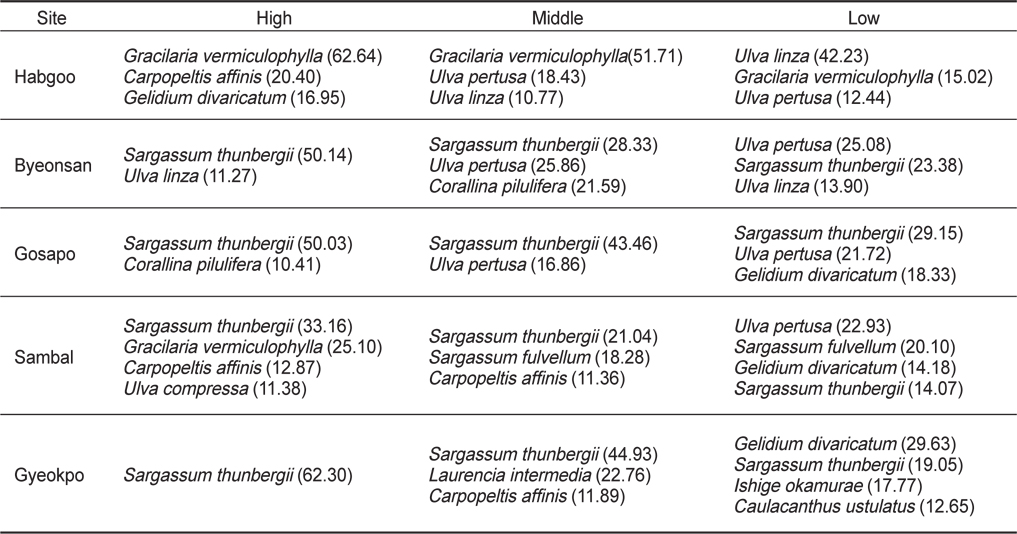
해조류의 중요도(IV)로 변산반도 조간대의 조위별 수직분포를 보면, 지충이(26.43)는 합구를 제외한 조사정점의 조간대 상·중·하부에서, 꼬시래기(38.61)는 합구(상·중·하부)와 삼발(상부)에서 우점하였고 잎파래(26.75)는 합구(하부)와 변산(상부)에서 주요종이었다(Table 3). 한편, 홍조류 까막살(20.40)은 합구의 상부, 작은구슬산호말(10.41)은 고사포의 상부, 참모자반(19.19)은 삼발의 중·하부,검은서실 (22.76)과 애기우뭇가사리(26.93)는 격포의 중 ·하부에서 번무하였다.
Vertical distribution of dominant seaweeds (IV>10) growing on the intertidal rocky shores at five study sites of Byeonsan Peninsula, Korea
해조류의 평균 생물량으로 산출한 우점도지수 (DI)는 변산에서 제 1, 2 우점종인 지충이(52.09 g dry wt/㎡)와 구멍갈파래(10.27 g/㎡)가 전체 해조류 생물량(140.77 g/㎡)의 44.30%를 차지하여 최소였고(0.44), 고사포에서는 지충이(54.52 g/㎡)와 구멍갈파래(2.93 g/㎡)가 생물량의 60.50% (총 생물량 94.96 g/㎡)를 차지하여 0.61로 최대였다(Table 4). 출현종수와 관련되는 풍도지수(R)는 44종이 출현한 삼발에서 최대(10.35)였고, 32종이 출현한 변산에서 6.27로 최소였다(Table 4). 출현종수와 생물량에 따라 변화하는 균등도지수는 출현종수가 적고 출현한 해조류의 생물량의 차이가 크지 않은 합구에서 최대였다. 풍도와 균등도에 의해 계산되는 종다양도지수(H’)는 삼발에서 가장 높은 값(1.85)을 보였으며 합구(1.71), 변산(1.65), 격포(1.52)와 고사포(1.49)의 순서로 나타났다.

Average biomass (g dry wt/㎡ ), percent cover (%) and various community indices of seaweeds at five study sites of Byeonsan Peninsula, Korea
해조상의 지역적 특성을 나타내는 C/P값은 변산반도 5개 정점에서 0.65였으며, 변산에서 최대(1.80), 삼발에서 최소(0.62)였다. R/P 값도 본 연구해역에서 2.4였고 변산에서 최대(3.80), 삼발에서 2.17로 최소였다(Table 4). (R+C)/P값의 경우는 변산 반도 5개 정점에서 3.0 이었으며, 정점별 2.92–5.60으로 C/P, R/P 값과 마찬가지로 변산에서 최대(5.60)였고 삼발에서 2.92로서 최소였다(Table 4).
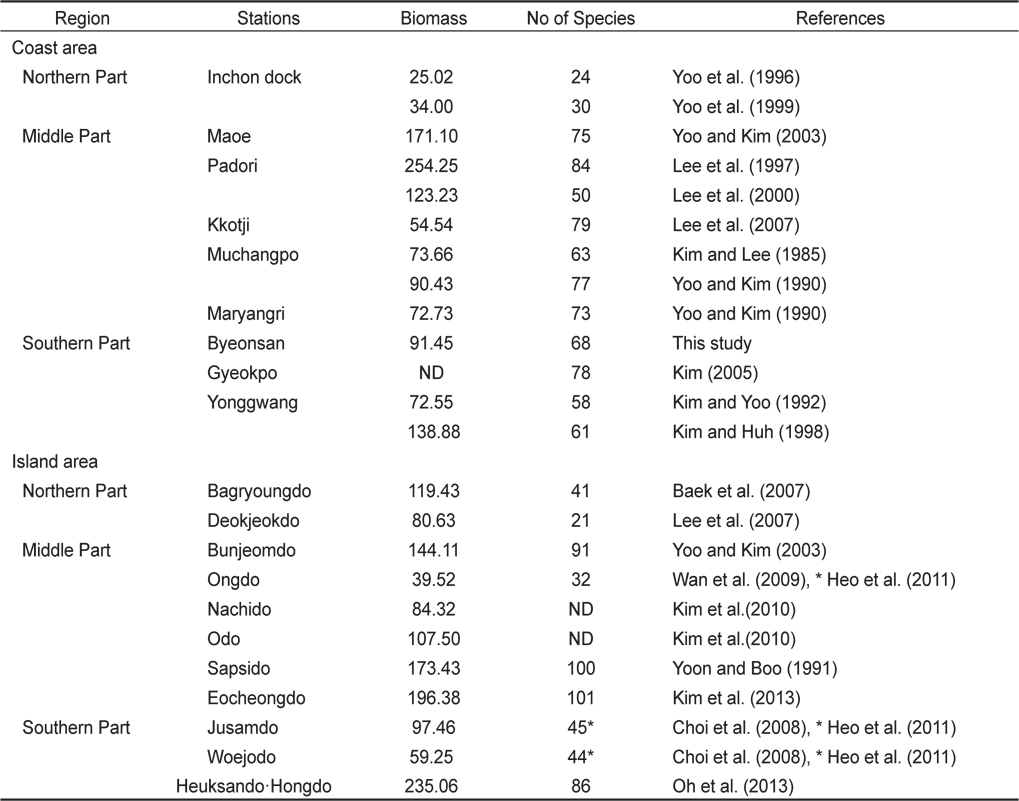
해조류의 분류군별 종조성과 출현종수(풍도)는 연안환경의 상태와 기후변화를 평가하는 생물학적인 지표자(biological indicator)로 활용된다(Wells et al., 2007). 본 연구에서 변산반도의 5개 정점에서 채집된 해조류는 68종이었고 정점별로는 18-51종으로 차이를 보였다. 서해안의 조간대에서 해조류 출현종은 북부해역(태안군 이북)에서 30종이하 (Yoo et al., 1996, 1999), 중부해역(변산 – 태안)에서 50–84종, 남부해역(신안 - 변산)에서 58–78종으로 태안군 인근에서 종다양성이 높게 나타났다(Table 5). 서해안의 도서 지역의 해조류 출현종수는 북부 해역에서 21–41종, 중부에서는 100–101종, 남부에서는 86종이 확인되었으며, 하계 조사만 실시된 도서에서 해조류 출현종수는 32–45종으로 매우 낮게 나타났다(Table 5).
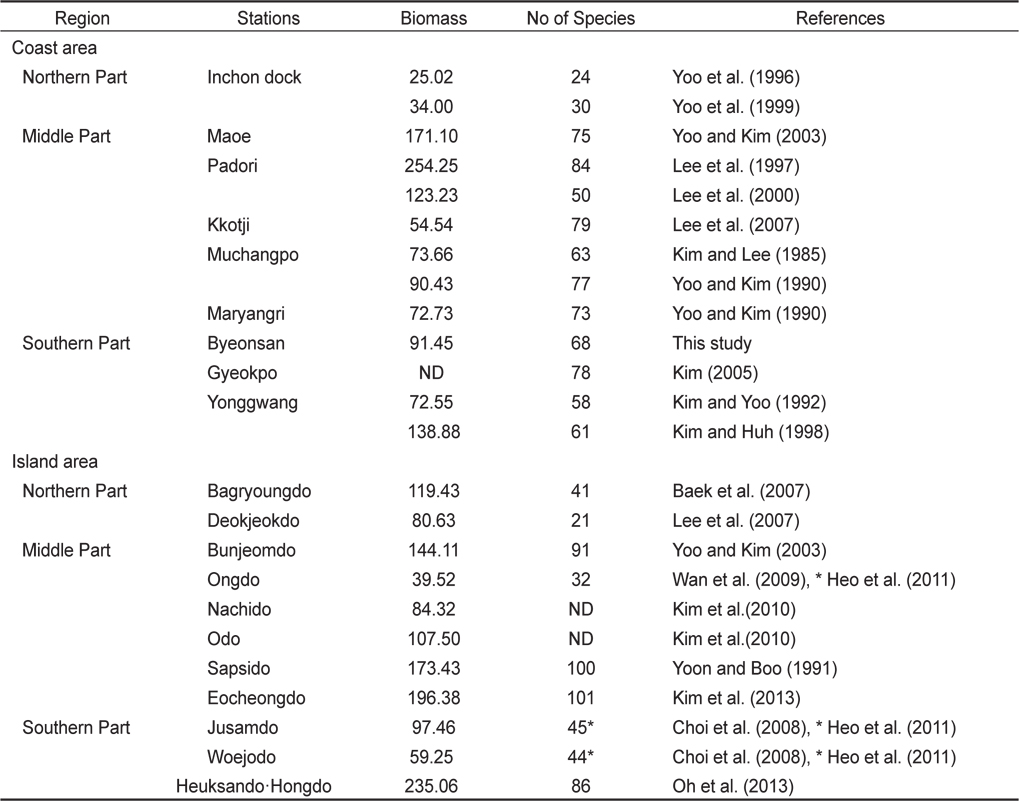
Annual seaweed biomass (g dry wt/㎡ ) and species richness at the intertidal zone of coastal and island areas along the Yellow Sea of Korea
Silva et al. (2012)은 브라질에서 관광객의 접근 허용해역과 금지해역을 비교해 본 결과, 해조류의 생물량, 출현종수, 종다양성은 관광객 접근 금지해역에서 높았고 우점도는 관광지에서 높게 나타났다고 하였다. 본 연구가 수행된 변산반도 연안의 해조류 종다양성은 백령도를 제외한 도서 지역에 비해 낮게 나타났으나, 태안과 유사하였고 영광에 비해 높게 나타났다. 하지만, 본 연구정점의 하나인 격포에서 해조류 출현종수가 78종에서 51종으로 약 10년 사이에 27종이 급감하였는데 종다양성 감소 원인이 격포 인근의 관광지 개발에 따른 부영양화 혹은 기후 변화와 관련이 있는지는 향후 좀더 지속적인 연구 수행을 통하여 밝혀야 할 과제이다(Kim, 2005).
서해안 중부해역인 변산반도 5개 정점의 조간대 해조류의 연 평균 생물량(건중량, g dry wt /㎡)은 91.45 g/㎡로서, 중부해역의 태안군 파도리(123.23–254.25 g/㎡)와 외해에 위치한 삽시도, 어청도와 흑산도·홍도에 비해 낮게 나타났으나 다른 지역과는 큰 차이가 나타나지 않았다(Table 5). 본 연구해역에서 지충이는 해조류 생물량의 47.89% 였으며, 고사포에서 생물량의 57.64%, 격포에서 56.70%를 차지하여 가장 우점하는 종으로 확인되었다(Table 2). 다른 연구결과에서도 지충이는 무창포에서 해조류 생물량의 57.83%, 마량리에서 56.88%, 그리고 주삼도에서 54.64%를 차지하여 서해안의 대표종으로 나타났다(Yoo and Kim, 1990; Lee et al., 1997; Lee et al., 2000; Choi et al., 2008). 또한, 최근에 녹조 대발생(green tide)을 유발하는 기회종인 갈파래류(
해조류의 분류군별 출현종수를 기준으로 해조류의 지리적 분포 특성을 나타내는 C/P값은 한대에서 아열대 해역에 걸쳐0.4–1.5범위의 값을 나타내고(Segawa, 1956), R/P값은 한대와 극지역에서 ≤1.0이하, 온대지역에서 1.0–4.0, 열대해역에서 ≥4.0 이상을 나타낸다(Feldmann, 1937). 또한, Cheney (1977)는 (R+C)/P값에 따라, 온대성 내지 한대성(< 3), 혼합성(3–6)과 열대성(> 6)의 해조상으로 구분한다. 본 연구해역인 변산반도에서는 C/P값이 0.65, R/P값은 2.35로 온대해역 해조상을 보였으며, (R+C)/P값은 3.00으로써 혼합성 해조상을 보였다. 우리나라 연안에서 분석된 해조류의 지리적 분포는 본 연구와 매우 유사하게 나타나고 있으나(Lee et al., 2007; Oh et al., 2013), C/P, R/P, 그리고 (R+C)/P값은 연안해역과 외해에 따라, 그리고 본 연구에서 보여준 것처럼 정점별로 값들이 커다란 차이를 보임으로써 이러한 값들의 사용 여부에 대한 심도 깊은 연구가 수행되어야 할 것으로 예견된다.
서해안 변산반도 5개 정점에서 연안생태계의 기초생산자인 해조류 종다양성과 생물량은 전반적으로 볼 때 다른 해역에 비해 크게 높지 않았으나, 합구 지역은 출현종수와 생물량이 매우 낮았으며, 나머지 정점에서는 비교적 양호하였다. 한편, 5개 정점에서 해조류 출현종수는 영양염과 퇴적물의 공급원인 만경강과 동진강 하구역에 있는 새만금 방조제에서 멀리 위치할수록 증가하는 경향을 보였고, 생물량은 변산(140.77g dry wt/㎡)과 격포(145.33g dry wt/㎡)에서 유사하였으나, 전체 생물량에 대한 갈파래류 비율은 새만금 방조제와 가까운 변산(11.44%)에서 격포(< 1%)에 비해 높게 나타났다. Wells et al. (2007)은 해조류의 종풍부도(species richness)는 환경이 바뀌지 않으면 거의 일정하기 때문에, 출현종수는 해조류 군집의 안정성을 반영하는 좋은 매개변수라고 하였다. 변산 반도의 5개 정점에서 출현종수, 생물량과 다양한 군집지수에서 가장 양호한 격포 해안에서 10여년 전에 수행된 연구결과와 비교해 볼 때 출현종수가 27종이 급감하는 것으로 확인되었다(Kim, 2005). 출현종수를 분류군별로 비교하면, 본 연구에서 갈조류 6종과 홍조류 21종이 감소하였고, 부영양화 해역에서 출현하는 녹조류인 대마디 말류(