The energy budget of the larvae of pandalid shrimp,
동물의 에너지수지에 관한 연구는 에너지 흐름을 이해하고 동물생태계의 기능을 파악하는데 있어 유용한 수단이 된다. 생태계는 복잡한 먹이사슬 또는 먹이그물과 같은 영양적 구조로 집단 간의 에너지흐름의 양에 밀접하게 관련되어 있다. 이러한 복잡한 먹이망 속에서 물질을 통한 에너지 흐름을 파악하기 위해서 에너지수지 분석은 각 생물의 영양대사 파악에 매우 중요하다(Klekowski and Duncan, 1975). 어느 한 생물의 개체에 유입되는 모든 에너지는 열로서 상실하거나, 다른 영양 단계로 전이된다. 생물학적 물질계는 물질전환에 따른 에너지 흐름에 의해서 체계화되고, 물질의 량과 질에 밀접하게 관련된다. 그러므로 에너지 흐름의 파악은 자연집단의 에너지수지를 분석하는 것이 필요하지만, 우선 개체의 생활사에 따른 에너지수지를 정확하게 분석하는 것이 매우 중요하다(Park, 1989).
갑각류의 에너지수지에 관해서는 자연과 실험실 내 집단을 대상으로 많은 연구가 진행되어 있으며(Crisp, 1974; Levine and Sulkin, 1979; Emmerson, 1984; Harms, 1987; Willows, 1987; Kim and Chin, 1991), 성장, 산소소비, 질소배설 및 섭식에 대한 에너지 함량을 파악하여 에너지수지 방식을 통해서 그 영양적 체계를 구성해 볼 수 있다(Clutter and Theilacker, 1971; Sameoto, 1976). 동물의 에너지수지에 관한 연구는 다른 영양 단계 간의 에너지 전이유형을 이해하는 총체적 지표가되며, 그 동물의 생산성과 생태적 효율을 예측하는 자료로서 유용하다(Vernberg and Vernverg, 1981).
물렁가시붉은새우(
본 연구는 물렁가시붉은새우 유생을 대상으로 성장, 탈피, 호흡, 섭취 및 에너지 함량을 측정하여 유생발달 동안 에너지수지를 분석하고 섭식과 성장에 따른 유생의 에너지 흐름의 경향을 파악함으로써 개체 및 개체군에 대한 생리적 기초 자료를 얻는데 그 목적이 있다.
초기 유생의 개체별 성장을 측정하기 위하여 수온 8±0.3℃ 하에서, 250 mL 용기(사육수, 200 mL)를 수용한 후 부화 직후 유생을 1마리씩 수용하여
유생의 산소소비량은 250 mL 산소병에 물렁가시붉은새우 단계별 유생을 5마리씩 수용하였고 수용직후와 24시간 후의 용존산소측정기(YSI 556 MPS, USA)로 용존산소를 측정하여 그 차이 값으로 정량하였다. 산소소비량 실험은 오전시간에 8℃사육수에 산소병을 담궈서 5회 반복으로 실시하여, 평균치로 개체당 단위시간당 산소소비량을 μLO2/ind/day으로 표시하였다. 산소소비량은 Gnaiger (1983)가 이용한 산소분자의 에너지 전환계수를 이용하였다(1 mgO2=3.38 cal). 대사에 쓰인 에너지는 4.71×10-3 cal/μLO2의 변환계수를 사용하여 계산하였다(Crisp, 1974). 개체의 산소소비량은 환경수의 산소 분압에 영향을 받기 때문에(Ghiretti, 1966; McLusky, 1973; Ikeda, 1974) 산소소비 측정이 끝날 때의 실험용기 내 용존산소량이 실험시작 시 산소량의 60.0% 이하로 떨어지지 않게 조절하였다.
유생의 섭취량 측정 시에는 250 mL의 용기에 물렁가시붉은새우 유생을 1마리씩 수용한 후 일정량의 계수된
각 유생단계 및 전체 유생발생에 대한 에너지 수지는 Standard IBP 방정식(Klekowski and Duncan, 1975)으로 나타내었다.
에 C는 섭취에너지, Pg는 체성장에 쓰인 에너지, Pe는 탈피체에 쓰인 에너지, R은 호흡에 쓰인 대사에너지, 그리고 U (뇨배설) 및 F (분배출) 에너지는 섭취에너지(C)와 동화에너지(A =Pg + Pe + R)간의 차로서 나타내었다(Dawirs, 1983).
수산동물의 동화효율 측정은 먹이섭취량에 대한 동화량을 백분율로 하여 구하였다. 물렁가시붉은새우의 유생에 대한 섭취, 성장, 탈피 및 호흡을 측정한 자료를 사용하여 다음 효율들을 계산하였다.
②식은 섭취량 중 동화된 양의 백분율을 나타낸다.
③식은 섭취량 중 성장으로 전환된 양의 백분율을 나타낸다.
④식은 동화량 중 성장으로 전환된 양의 백분율을 나타낸다.
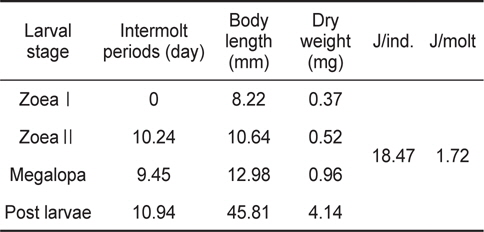
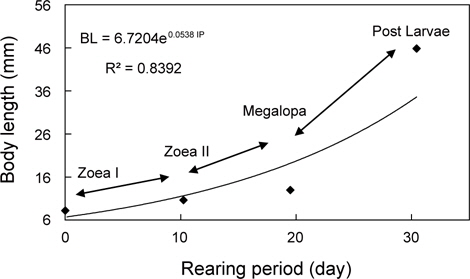
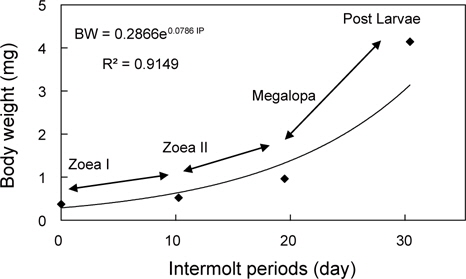
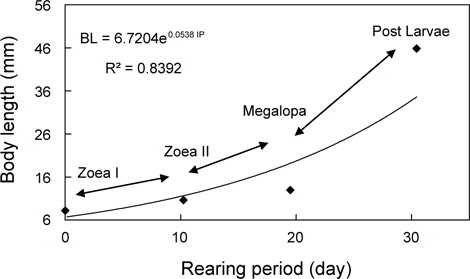
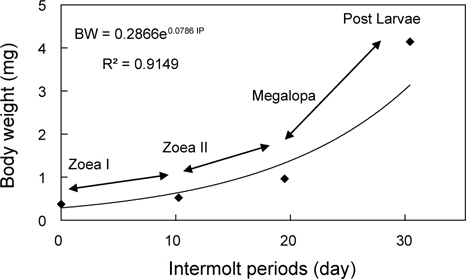
물렁가시붉은새우의 부화 후 경과시간에 따른 유생의 체장, 건조중량 및 유생과 탈피체의 에너지함량은 Table 1에 나타내었다. ZoeaⅠ기에서 post larvae기까지의 체장성장과 체중성장은 지수적인 증가를 보였으며, 각각 BL=6.7204e0.0538 IP (R²=0.8392)와 BW=0.2866e0.0786 IP (R²=0.9149) 으로 나타났다(Fig. 1과 2).
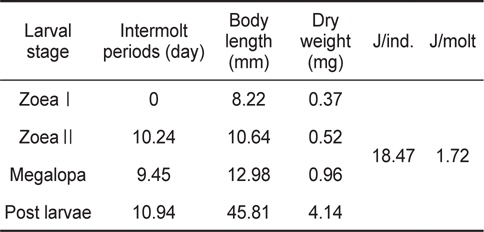
The intermolt periods (day), body length (mm), body weight (mg) and total energy contents (J/ind. and J/molt) up to post larvae stage of Pandalopsis japonica larvae for different larval stages
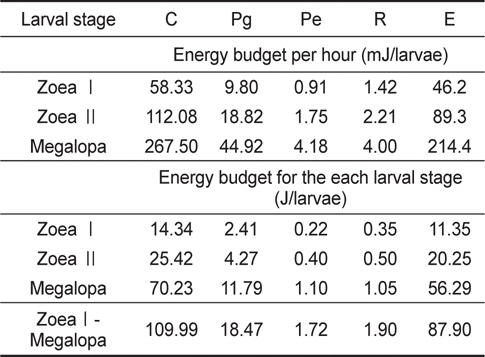
성장에 사용된 에너지 즉, 생체 에너지 함량은 J/ind.로 산출하였다. ZoeaⅠ기에서 post larvae기까지 변태하는 동안 megalopa기의 생체에너지는 18.47 J의 에너지를 체성장에 사용하였으며, 성장에 수반되어 zoeaⅠ기에서 post larvae기까지 탈피하는 동안 탈피체 에너지로 사용한 것은 1.72 J이었다(Table 1).
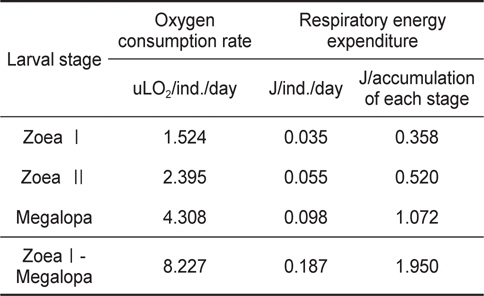
물렁가시붉은새우 유생기의 산소소비량은 zoeaⅠ기의 1.524 uLO2/ind./day에서 megalopa기의 4.308 uLO2/ind,/day로 유생발달에 따라 증가하였다(Table 2).
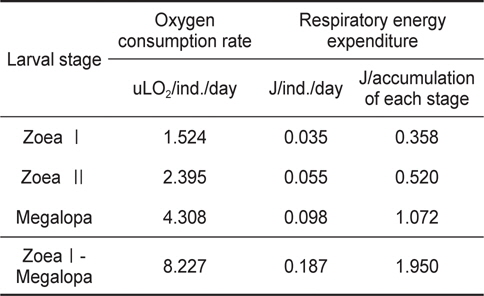
Individual oxygen consumption rate and respiratory energy expenditure of Pandalopsis japonica larvae on the different larval stages
Table 2에 나타낸 일간대사에너지를 사용하여 Table 1에 표시된 각 유생단계별 경과시간에 따른 대사에너지를 계산한 결과 zoeaⅠ기 기간 동안에는 0.358 J, zoeaⅡ기는 0.520 J, megalopa기 기간 동안에는 1.072 J을 각각 소비하여 유생기간 동안 소비된 총 산소소비에너지는 1.950 J이었다 (Table 2).
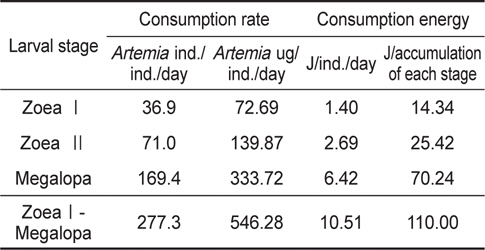
물렁가시붉은새우의 zoea 유생들에 의해 섭취된 일간섭취량과 일간섭취에너지를 Table 3에 나타내었다. ZoeaⅠ기의 36.9
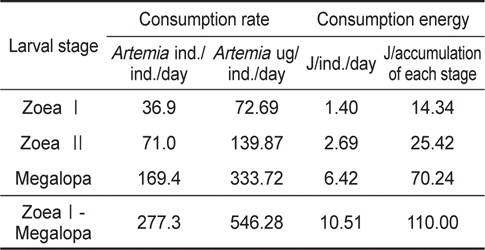
Consumption rate and consumption energy of Pandalopsis japonica larvae on the different larval stages
성장에 따른 전 유생기 동안의 유생의 에너지함량(Pg) 및 탈피체의 에너지함량(Pe)은 Table 1에서 제시한 결과를 사용하였으며, 호흡 대사에너지(R)는 Table 2의 자료로부터 계산하였다. 그리고 섭취에너지(C)는 Table 3의 일간 섭취량과 일간 섭취에너지로부터 계산하였다. 뇨배설(U) 및 분배출(F) 에너지는 섭취에너지(C)와 동화에너지(A)의 차이로서 계산하였다. 전 유생기 동안의 누적에너지수지를 Table 4에 나타내었다. ZoeaⅠ기에서 megalopa기까지의 총 누적섭취에너지는 109.99 J이었으며, 이것은 성장에 18.47 J, 탈피체 손실에 1.72 J 그리고 호흡에 1.90 J의 에너지를 사용하며 뇨와 분으로 87.90 J의 에너지가 배설과 배출이 이루어지는 것으로 조사되었다.
[Table 4.] Energy budgets of Pandalopsis japonica larvae on the different larval stages
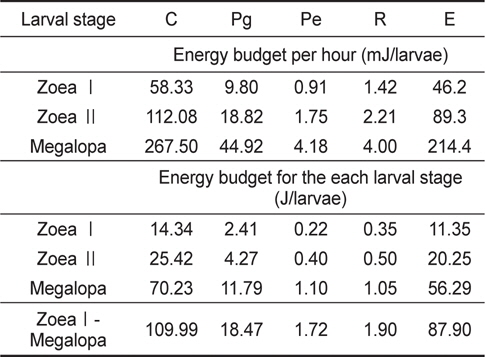
Energy budgets of Pandalopsis japonica larvae on the different larval stages
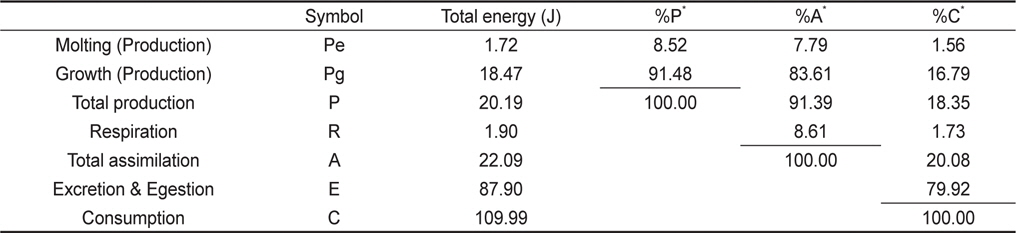
물렁가시붉은새우 유생의 섭취, 성장, 탈피 및 호흡을 측정한 자료로부터 생태효율을 Table 5에 나타내었다. 유생의 동화효율은 20.08%이었으며, 총 성장효율(K1)은 18.35%, 순 성장효율(K2)은 91.39%로 나타났다. 각 에너지수지 구성요소들의 섭취에너지에 대한 백분율은 성장이 18.36%, 탈피체가 1.56%, 대사에너지가 1.73% 그리고 뇨와 분으로 79.92%이었다. ZoeaⅠ기부터 megalopa기까지의 총 동화에너지는 22.09 J이었으며, 각 에너지수지 구성 요소들의 동화에너지에 대한 백분율은 성장이 83.61%, 탈피체가 7.79% 그리고 대사에너지가 8.60%이었다. 또한 zoeaⅠ기에서 megalopa기까지의 총 생산 에너지는 총 동화에너지의 91.40%, 총섭취에너지의 18.36%인 20.19 J 이었다.
[Table 5.] Accumulative energy budget of Pandalopsis japonica larvae for whole larval stages
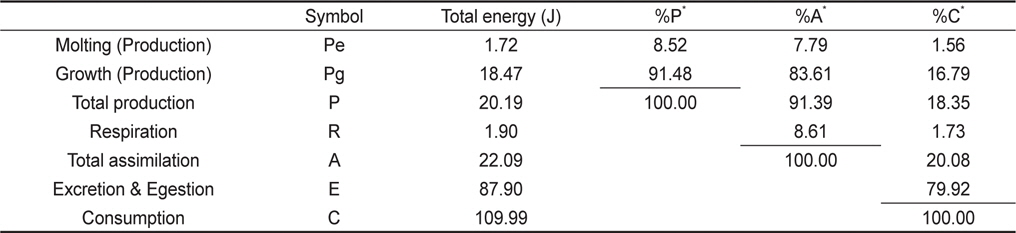
Accumulative energy budget of Pandalopsis japonica larvae for whole larval stages
해산동물의 에너지 흐름을 파악하기 위해서는 그 생물의 생활사를 정확하게 조사하는 것이 선행되어야 한다. 갑각류의 성장은 환경에 따라 다양한 반응을 보이지만(Hartnoll and Dalley, 1981), 특히 섭취, 성장 및 대사율은 생육단계별로 많은 변동을 보이는 것으로 알려져 있다(Reeve, 1969; Hiroko et al., 1990). 십각목 갑각류 유생의 성장은 일반적으로 체장성장에 따른 체중성장이 지수적으로 나타낸다(Mootz and Epifanio, 1974; Logan and Epifanio, 1978; Johns and Pechenik, 1980; Stephenson and Knight, 1980; Dawirs, 1983; Park, 1989). 본 연구에서도 물렁가시붉은새우의 유생은 타 어종과 동일한 지수적 체성장을 보였다. 이와 같은 결과는 Decapoda의
물렁가시붉은새우 유생의 산소소비율은 유생발육이 진행될수록 개체당 소비율의 증가를 보인다. 이는
물렁가시붉은새우 유생의 동화효율은 20.08%로, 본 연구와 같은 유생의 동화효율은 새우류인
물렁가시붉은새우의 총 성장효율(K1)은 18.35%로서 본 연구종과 비슷한 크기의 새우류인 보리새우과의
결과적으로 생태생리학적 생태효율의 차이는 개체의 크기와 갑각류 내 종류에 따라 많은 차이를 보이는 것으로 판단된다. 또한 환경인자들의 변화는 따른 생물적응은 각기 다른 생화학적, 생리학적 그리고 행동 정도에 따라 다르게 나타나는 종 특이적 항상성(homeostasis) 조절 메카니즘에 의한 복합적인 반응(Spanopoulos-Hernandez et al. 2005)에서 오는 것으로 판단 된다.