The feeding habits of
벌레문치(
등가시치과에 관한 연구는 러시아 Primor&e 해역과 타타르 해협에서 벌레문치의 체급별 식성연구(Saveliev et al., 2011, 2012)를 비롯하여 등가시치류와 베링해에 서식하는 라리벌레문치(
동해안의 해저에는 가시베도라치, 주먹물수배기, 청자갈치, 분홍꼼치(
벌레문치는 2011년 3월부터 2012년 12월까지 강원도 동해연안의 100-800 m 수심에서 동해구기선저인망과 자망어업에 의해 어획된 것을 위판장에서 매월 채집하였다(Fig. 1). 벌레문치의 위내용물은 어체 생물측정 후 복부를 절개하고 위를 분리하여 바로 분석을 실시하였고 위내용물 중에서 발견된 먹이생물은 육안 또는 해부현미경을 사용하여 관찰한 후 Kim et al. (2005), Hong (2006), NFRDI (2001, 2005) 등의 도감을 이용하여 동정하였다.
위내용물의 먹이생물은 종별로 개체수를 계수하였고 각 먹이 생물의 크기는 0.1 mm 단위까지, 중량은 습중량으로 0.01 g 단위까지 측정하였다. 먹이생물 중요성의 척도에 대한 지수는 개체수비(Number of individuals, %N), 습중량비(Wet weight, %W) 및 출현빈도(Frequency of occurrence, %F)들을 고려한 Pinkas et al. (1971)의 상대중요성지수(IRI, Index of Relative Importance)를 이용하였다.
전장에 따른 벌레문치의 먹이생물 변화를 파악하기 위해서 5 cm간격으로 구분하여 각 전장별 먹이생물을 관찰하였으며, 또한 계절에 따른 변동을 파악하기 위하여 월별 먹이생물 조성을 분석하였다.
벌레문치 위내용물에서 관찰된 먹이생물의 다양도과 우점도를 파악하기 위해 각 먹이생물군의 풍도(
ni = 각각의 전장별(월별)에서 출현한 i 번째 종의 개체수
Simpson (1949)의 종우점도지수(
위내용에서 출현한 먹이생물 조성의 유사성을 파악하기 위해 전장별·월별로 3회 이상 출현한 먹이생물의 종수(C
BC
각각의 전장별(월별)로 분석된 유사도를 바탕으로 전장별(월별) 유사성의 차이를 분석하기 위하여 수지도(dendrogram)를 작성하고, 다차원척도분석(MDS, Multi Dimensional Scaling)을 실시하여 2차원 공간에서의 군집구조의 차이를 파악하기 위해 집괴분석(cluster analysis)을 실시하였으며, 그룹간 연결 방식에는 complete linkage 방식을 사용하였다. 전장·월별로 분리된 그룹 사이의 먹이생물의 유사성 차이를 유발하는 종을 분석하고 이들이 먹이생물 유사성의 차이에 기여하는 정도를 분석 하기 위하여 SIMPER 분석을 실시하였으며, 이들의 분석에는 PRIMER V5.0을 사용하여 실시하였다(Clarke and Warwick, 2001).
벌레문치의 산란시기와 체중에 따른 위내용물 중량간의 관계를 알아보기 위하여 벌레문치의 전중(TW, Total Weight)에 대한 생식소 중량(GW, Gonad Weight) 비율인 생식소 중량지수(GI, Gonad Index)와 위내용물 중량(SCW, Stomach Content Weight) 비율인 위내용물 중량지수(SCI, Stomach Contents Index; Watanabe et al., 2004)를 비교하였다.
GI = GW/TW×100
SCI=SCW/TW×100
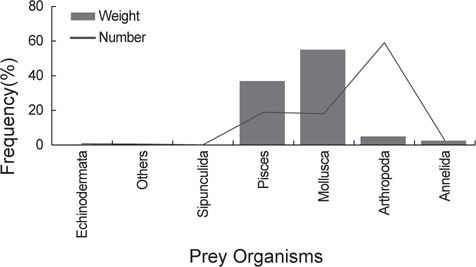
벌레문치는 총 722개체를 분석하였으며 위내용물이 없었던 개체는 총 368개체로 50.9%의 공복율을 보였으며 위내용물이 발견된 354개체의 위내용물을 분석하였다. 벌레문치의 위내용물에서 발견된 먹이생물은 어류(Pisces) 17종, 연체동물(Mollusca) 9종, 절지동물(Arthropoda) 13종, 환형동물(Annelida) 3종, 극피동물(Echinodermata) 1종, 성구동물(Sipunculida) 1종으로 총 44종이 발견되었다.
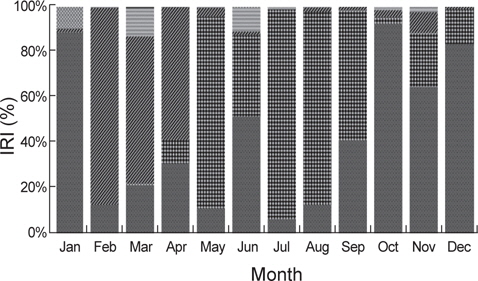
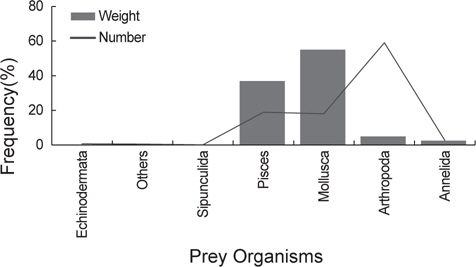
출현 먹이생물의 총 개체수비는 절지동물이 59.0%로 가장 높았고 어류와 연체동물이 각각 18.9%, 18.0%로 나타났다. 생체량비는 연체동물이 55.0%로 가장 많은 비중을 차지하였고 다음으로 어류가 37.0%이었다. 개체수비에서는 상대적으로 크기가 작은 절지동물, 환형동물 등이 차지하는 비율이 높았고 생체량비에서는 크기가 큰 연체동물과 어류의 비율이 높게 나타났다(Fig. 2).
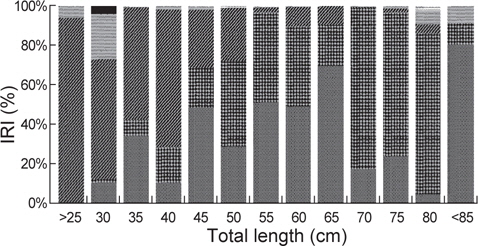
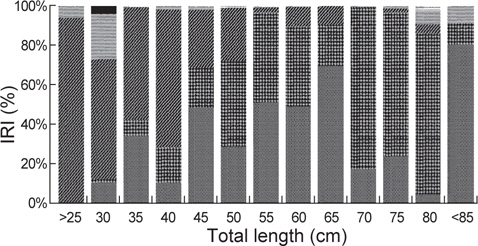
벌레문치 전장(25 cm미만-80 cm이상)을 13개 계급으로 구분 하여 위내용물의 먹이생물을 관찰한 결과, 16-40 cm의 소형군에서 절지동물이 상대중요성지수에서 56.8-94.0% 범위로 우점하였으며, 전장이 작을수록 단각류와 난바다곤쟁이류(Euphausiacea sp.)를 섭식하고 전장이 커지면서 진흙새우를 주로 섭식하는 것으로 나타났다. 전장 41-65 cm에서 주요 먹이생물은 어류와 연체동물로서 어류는 상대중요성지수에서 28.9-69.7%의 범위를 보였는데, 전장 41-55 cm에서는 큰눈등가시치(
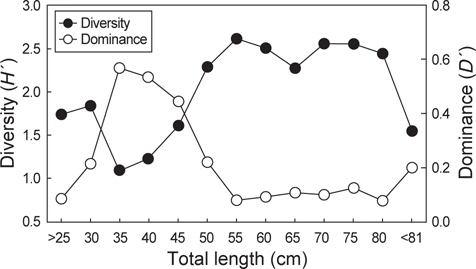
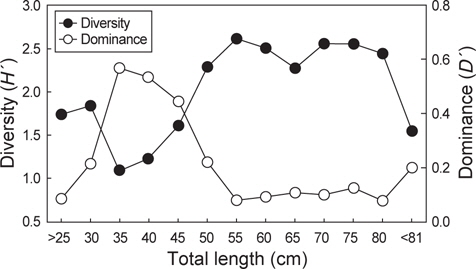
벌레문치가 섭이한 먹이생물의 종수와 개체수에 대한 종다양도와 종우점도를 전장별로 분석한 결과, 종다양도는 51-55 cm 에서 2.61로 가장 높았으며 66-70 cm와 71-75 cm 전장군에서 각각 2.55와 2.54로 다른 전장군에 비해 비교적 높게 나타났다. 반면, 31-35 cm, 36-40 cm, 41-45 cm 전장군에서는 각각 1.09, 1.22, 1.60으로 나타나 51 cm이상의 전장군에 비해 종다양도가 낮은 값으로 나타났다.
종우점도는 31-35 cm에서 0.57로 가장 높았고 36-40 cm, 41-45 cm의 전장군에서 각각 0.53과 0.44로 다른 전장군에 비해 비교적 높았다. 반면, 51-80 cm의 전장 범위에서는 0.10-0.13의 낮은 우점도를 보였으며 종우점도는 종다양도의 변동과는 대체적으로 반대 경향을 보였다(Fig. 4).
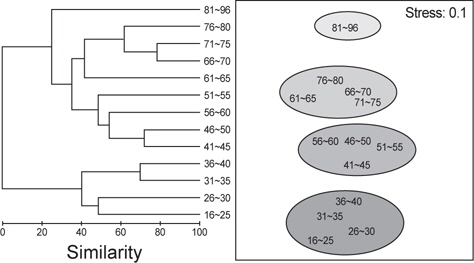
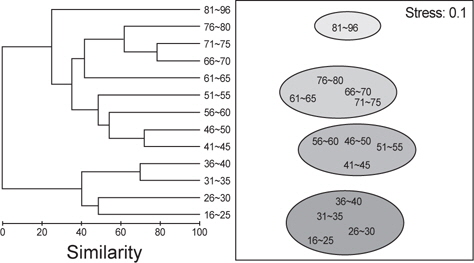
벌레문치의 크기에 따른 먹이생물조성의 유사성을 파악하기 위해 13개의 크기군에 대한 군집분석을 실시한 결과 4개의 그룹(Ⅰ: 16-40 cm, Ⅱ : 41-65 cm, Ⅲ : 66-80 cm, Ⅳ : 81-96 cm)으로 구분되었다. 그룹Ⅰ에서 가장 많이 섭이한 먹이는 절지동물로 상대중요성지수에서 평균 70.2%를 차지하였는데 그 중에서 단각류(35.9%)와 진흙새우(17.0%)가 가장 중요한 먹이원으로 나타났다. 그룹Ⅱ의 주요 먹이생물은 어류와 연체동물로 각각 평균 37.4%와 34.1%를 차지하였는데, 어류 중에서는 청자갈치(13.6%)와 멸치(7.3%)가 주로 나타났고, 연체동물 중에서는 매오징어(16.8%)와 대문어(6.6%)가 주로 관찰되었다. 그룹Ⅲ에서 주요 먹이생물은 연체동물이 평균 78.6%를 차지할 정 도로 극우점하는 경향을 보였는데 이 중에서 갈고리흰오징어(45.7%)와 매오징어(21.2%)를 주로 섭식하는 것으로 나타났으며, 그룹Ⅳ에서 주요 먹이생물은 어류로 상대먹이중요성지수에서 80.5%를 차지하였다(Fig. 5). 벌레문치의 성숙체장은 60 cm 이상으로 전장별 먹이생물의 유사성은 성숙체장보다 작은 개체들에서는 절지동물이나 크기가 작은 연체동물과 어류를 주로 섭이하였고 성숙체장 이상의 개체들은 크기가 큰 연체동물과 어류를 주로 섭식하는 것으로 나타났다.
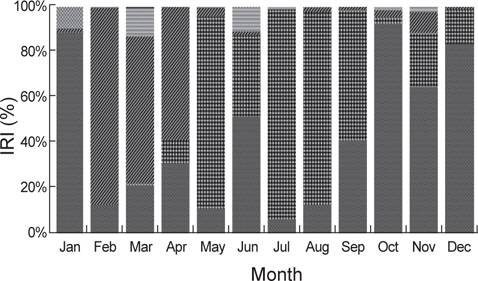
벌레문치의 월별 위내용물 조성의 상대중요성지수 변화를 살펴보면, 1월에는 어류가 88.3%의 높은 비율을 차지하였고 2월부터 4월까지는 절지동물이 각각 86.3%, 64.6%, 58.3%의 비율을 차지하였는데 주로 단각류와 진흙새우를 섭이하였다. 5월에는 절지동물의 비율이 감소한 반면, 연체동물의 비율이 83.9%로 높게 차지하였고 주로 매오징어를 섭이하였다. 6월에는 어류와 함께 연체동물의 비율이 각각 51.6%와 35.7%를 차지하며 중요한 먹이생물로 나타났으며, 7월부터 9월까지는 연체동물이 각각 92.1%, 84.5%, 57.1%의 높은 비율을 차지하였고 갈 고리흰오징어와 매오징어, 살오징어를 주로 섭이하였다. 10월부터는 연체동물의 비율이 감소하기 시작하고 어류가 92.1%의 높은 비율을 차지하였으며 주로 청자갈치를 섭이하였다. 11월과 12월에도 어류가 각각 64.6%와 83.2%의 비율을 차지하였고 먹갈치(
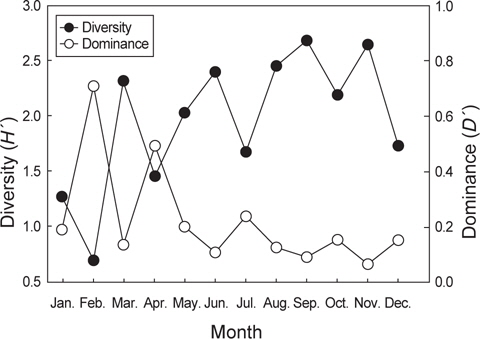
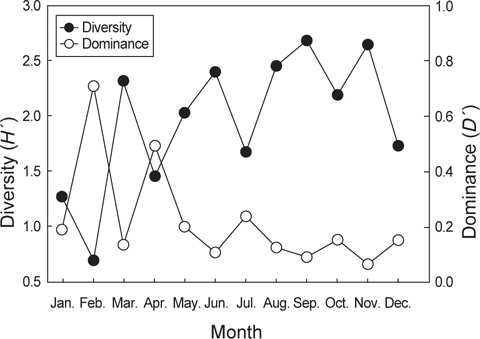
먹이생물의 종수와 개체수를 바탕으로 먹이생물의 종다양도와 종우점도를 월별로 분석한 결과, 종다양도는 가을철인 9월과 11월에 각각 2.68과 2.65로 가장 높게 나타났다. 9월에 출현한 종수는 총 23종으로 어류와 연체동물을 주로 섭식하는 것으로 나타났으며 어류에서는 멸치와 청자갈치를 연체동물에서는 매오징어, 살오징어를 주로 섭이하였다. 11월에 출현한 종수는 총 19종으로 어류, 연체동물과 절지동물을 고루 섭식하는 것으로 나타났는데 어류에서는 비교적 크기가 큰 먹갈치와 칠성갈치를 주로 섭이했고, 연체동물에서는 갈고리흰오징어와 좀귀오징어를 절지동물에서는 새우류를 주로 섭이했다. 반면 1월과 2월의 종다양도는 각각 1.28과 0.70으로 낮은 값을 보였으며 1월에는 4종으로 종수가 적어 종다양도가 낮았던 반면 2월에는 16종이 출현하였지만 먹이생물이 난바다곤쟁이류에 대부분 집중되어 있어 종다양도가 낮게 나타났다.
종우점도는 2월과 4월에 0.71과 0.50으로 가장 높았으며 먹이생물 중에서는 난바다곤쟁이류의 비율이 가장 높았다. 9월과 11월의 종우점도는 0.09와 0.07로 가장 낮았는데 9월과 11월은 종수도 높고 다양한 생물을 섭이하여 먹이에 대한 편중이 낮은 것으로 나타났다(Fig. 7).
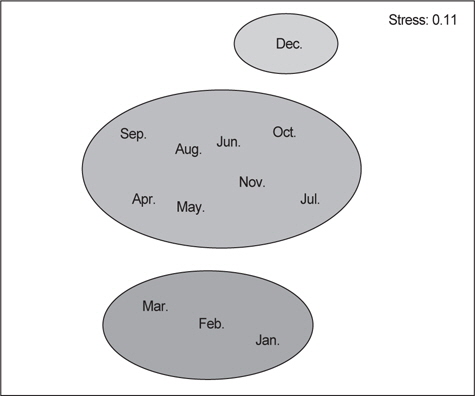
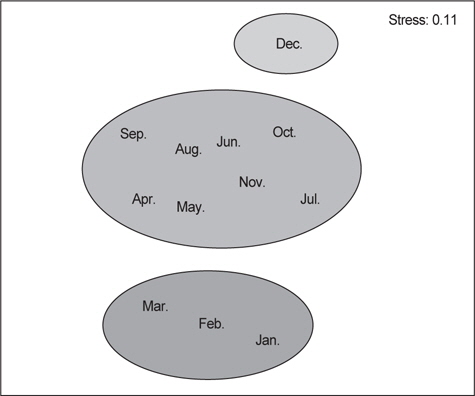
벌레문치의 월별 먹이생물의 유사성을 파악하기 위해 군집분석을 실시한 결과 3개의 그룹(I: 1-3월, II : 4-11월, III : 12월)으로 구분되었다.
I 그룹은 수온이 비교적 낮은 겨울철로 전장의 크기가 비교적 작은 개체들과 산란이 가까워진 큰 개체들이 유사한 수심에서 출현한 시기로 크기가 큰 개체들은 대부분 공위를 나타냈고, 크기가 작았던 개체들은 절지동물을 주로 섭이하는 것으로 나타났다. 절지동물의 상대성중요성지수를 살펴보면 51.0%로서 다른 먹이생물에 비해 매우 우점하는 것으로 나타났으며, 그 중 난바다곤쟁이류와 단각류가 가장 높은 비율을 보였다. II 그룹은 봄부터 초가을에 해당하는 시기로 대부분의 개체들은 35-70 cm 이었고 주요 먹이생물은 상대중요성지수가 48.7%인 연체동물이었으며 그 중 매오징어와 살오징어를 주로 섭이하였다. 또한, 어류의 상대중요성지수는 38.9%로서 겨울에 비해 상대적으로 먹이비율이 증가한 것으로 나타났다. III 그룹은 벌레문치의 산란이 시작되는 12월로서 대부분의 개체들이 70-85 cm 전장범위를 보이며 작은 개체들은 거의 출현하지 않는 특징을 보였다. III 그룹의 상대중요성지수에서는 어류가 83.2%로서 먹갈치와 청자갈치를 주로 섭이하였고, 연체동물의 상대중요성 지수는 15.3%로 상대적으로 섭이율이 낮았으며 갈고리흰오징어와 대문어를 주로 섭이하였다(Fig. 8).
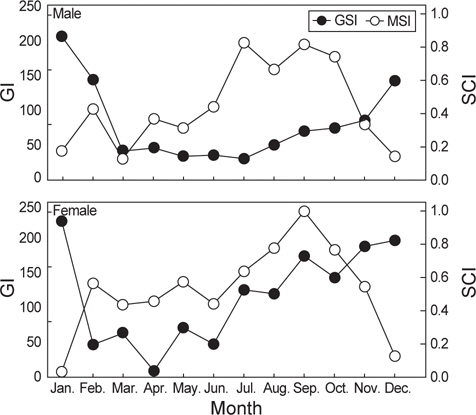
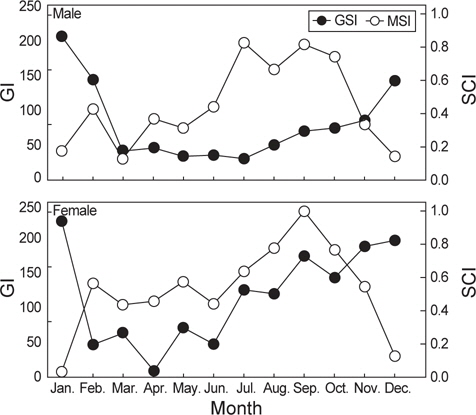
암컷의 생식소 중량지수의 월 변화를 살펴보면 미숙 시기인 2-6월까지는 평균 46.8의 값을 보이다가 중숙시기인 7-10월에 평균 139.6으로 증가하였으며 완숙기인 11-1월에 평균 201.7로 가장 높게 나타났다(Fig. 9). 미숙과 중숙기에는 생식소 중량지수가 증가함에 따라 위내용물 중량지수도 증가하는 경향을 보이다가 생식소 중량지수가 가장 높은 값을 보이는 완숙기에는 감소하는 것으로 나타났다. 또한 생식소 중량지수의 변화에 따른 공복 개체의 비율을 살펴보면 미숙기에는 13.4%, 중숙기에는 7.1%, 완숙기에는 79.5%의 공복율을 보였다(Fig. 9).
수컷도 암컷과 유사한 경향을 보였는데, 생식소 중량지수는 미숙기인 3-7월까지는 평균 5.2의 값을 보이다가 중숙기인 8-10월에는 평균 9.0의 값을 보였고, 완숙기인 12-2월에 평균 20.1의 가장 높은 값을 보였다. 수컷의 위내용물 중량지수 또한 미숙과 중숙기 까지는 생식소중량지수가 증가함에 따라 증가하는 경향을 보이다가 완숙기가 되면서 위내용물 중량지수가 감소하는 경향으로 나타났다. 생식소 중량지수의 변화에 따른 공복 개체의 비율은 미숙기에는 22.4%, 중숙기에는 10.4%, 완숙기에는 67.2%로서 암컷과 수컷 모두 완숙기가 되면서 공복율 이 크게 증가하는 경향을 보였다(Fig. 9).
본 연구에서 벌레문치 시료 중 위내용물이 전혀 발견되지 않은 개체는 368개체로 전체 출현 개체의 50.9%의 공복율을 보였다. 러시아 Primor'e 해역에서 벌레문치의 식성에 대한 연구에 의하면 40 cm이하, 41-50 cm와 51-60 cm의 전장에서 각각 47.1%, 46.7%와 41.2%로 상대적으로 낮은 반면 전장 61-70 cm와 70 cm이상에서는 55.6%와 50.0%로서 본 연구와 유사하거나 상대적으로 높은 공복율을 보였다(Saveliev et al., 2012). 우리나라 해역에서 조사된 저어류의 공복율을 보면 참가자미(
본 연구에서도 많은 어식성 어류에서 나타나는 공식현상 (Cannibalism)을 보였는데, 41-45 cm에서 1개체(10 cm), 46- 50 cm에서 2개체(12.1 cm, 13.6 cm), 51-55 cm에서 3개체(9.8- 10.6 cm), 71-75 cm에서 1개체(36.4 cm)로 총 7개체가 발견되었다. 일본 타타르 해역에서 조사한 벌레문치에서는 전장 40 cm에서 1개체(전장 11 cm)가 출현한 것으로 나타났지만(Saveliev et al., 2012), 황해에서 조사된 벌레문치 연구에서는 공식 현상이 나타나지 않았다(Jin et al., 2010). 우리나라 해역에 분포하는 어류로는 가시망둑(Huh and Kwak, 1998), 갈치(Huh, 1999), 황아귀(Baeck and Huh, 2003)와 꼬치고기(Baeck and Huh, 2004) 등에서 공식현상이 나타나는 것으로 보고하였다. 공식현상을 보이는 종들의 경우 벌레문치처럼 입이 큰 경우 어릴 때에는 절지동물과 환형동물과 같이 작은 크기의 먹이를 섭이하다가 성장하면서 입이 커지면 연체동물과 어류를 섭식하게 되면서 공식현상을 보이는 것으로 여겨지는데, 이와 같은 개체들은 대부분 일정 분류군을 선택하기 보다 먹이가 발견되는 대로 섭식하는 특성을 가진 것으로 판단된다.
벌레문치의 크기에 따른 먹이생물의 변화를 보면, 전장16-40 cm에서는 단각류, 난바다곤쟁이류, 새우류 등과 같은 소형 절지동물을 주로 섭이하는 반면, 전장 41-65 cm에서는 크기가 상대적으로 작은 매오징어와 살오징어, 큰눈등가시치와 가시베도라치를 주로 섭이하는 것으로 나타났으며 전장 66-80 cm의 큰 어체에서는 상대적으로 크기가 큰 갈고리흰오징어를 섭이하는 특징을 보였다. 벌레문치는 체장이 증가함에 따라 절지동물과 같이 크기가 작은 생물을 많이 섭이하는 특성에서 연체동물과 어류와 같이 크기가 큰 개체를 적게 섭이하는 먹이습성으로 바뀌는 경향을 보였는데, 이러한 경향은 입이 크고 강한 포식성을 보이는 갈치(Huh, 1999), 삼치(Huh et al., 2006), 전갱이(Cha, 1991), 황아귀(Cha et al., 1997), 게르치류(
벌레문치의 크기에 따른 먹이생물의 종다양도와 종우점도를 보면, 전장 16-40 cm에서 크기가 작은 난바다곤쟁이류, 새우류들을 많이 섭이하는 특성 때문에 종다양도가 1.47로 낮았고 종우점도는 0.35로 높게 나타났지만, 40 cm이상 크기에서는 절지동물의 먹이섭이 비율이 감소하고 갈고리흰오징어, 살오징어, 매오징어 등의 연체동물과 멸치, 줄가시횟대, 청자갈치 등의 어류와 같이 다양한 먹이생물을 소량으로 섭이하여 종다양도는 평균 2.26으로 증가된 반면 한 종에 대한 섭이의 편중이 감소하면서 종우점도는 평균 0.17로 낮아져 16-40 cm미만의 개 체와는 반대되는 경향을 보였다. Saveliev et al. (2011)의 연구에서도 크기가 작은 시기에는 많은 개체의 자주새우(
벌레문치의 월별 먹이생물의 차이를 분석한 결과 2-6월에 난바다곤쟁이류와 단각류를 많이 섭이하는 것으로 나타났는데, 이 시기는 난바다곤쟁이류의 산란기로서 여름과 초겨울(8-12월)동안 성장이 멈추었다가 산란을 위해 성장을 다시 시작하는 시기(Iguchi et al., 1993)로서 개체수와 생체량이 증가할 뿐만아니라 군집을 형성하여 수직 일주운동(Daily migration)을 하기 때문에 벌레문치 등 다양한 어류들의 먹이로 이용된다(Sancho et al., 2006; Balanov et al., 2006; Saveliev et al., 2011). 1-3월에 산란 참여하는 큰 개체는 대부분 공복상태이거나 크기가 큰 어류를 섭취한 반면 출현비율이 높은 40 cm미만의 개체는 난바다곤쟁이, 단각류와 같은 소형갑각류와 환형동물을 주로 섭식하는 것으로 나타나 상대적으로 절지동물이 중요한 먹이생물이었다. 봄과 여름철인 4-6월에는 매오징어, 살오징어에 대한 섭이비율이 높았는데, 이 시기는 벌레문치가 500 m 정도의 깊은 수심에서 200 m내외의 보다 낮은 수심으로 이동하면서 먹이활동을 활발하게 하는 시기로 동해안의 수심 200-300 m의 저층에서 무리지어 서식하는 매오징어와 비교적 크기가 작은 살오징어를 많이 섭이하는 경향을 보였다(MMAF and NFRDI, 2005; NFRDI, 2005). 7-9월에는 60 cm 이상의 큰 개체들이 산란을 준비하기 위해 왕성하게 먹이활동을 하는 시기로 수심 300-500 m 내외에서 서식밀도가 높은 것으로 알려진 갈고리흰오징어(Sohn et al., 2010)에 대한 섭이활동이 활발했고 겨울철이 되면서 벌레문치는 산란을 위해 500 m 전후의 깊은 수심으로 이동하면서 주로 어류를 섭이하는 것으로 알려져 있다(Park et al., 2007; Sohn et al., 2010; Saveliev et al., 2011; Balanov et al., 2011). 본 조사에서도 12월에 채집된 개체들은 대부분 70 cm상의 큰 개체들이었으며 주로 먹갈치와 청자갈치와 같이 크기가 큰 어류를 주로 섭이하는 것으로 나타났다. 본 조사에서 월별로 벌레문치의 먹이원이 다르게 나타난 것은 선택적으로 먹이원을 섭이하는 습성이라기 보다 성장 및 산란 등 생활사에 따라 서식지 이동을 하면서 먹이생물을 섭이하기 때문인 것으로 판단된다.
벌레문치의 주 산란기는 12-2월로서 생식소 크기가 급격하게 커지게 되며 이 시기 벌레문치의 공위율은 증가하고 산란이 끝나는 시기와 미숙·중숙기가 되면서 공위율이 감소하는 것으로 나타났다. 위내용물 중량지수는 암컷과 수컷의 성숙시기인 11-2월이 되면 가장 낮은 값을 보여 생식소 중량지수와 위내용물 중량지수는 서로 반대되는 경향을 보였다. 일반적으로 어류들은 산란기가 되면 생식소가 커지면서 복강에 생식소가 차지하는 비중이 커지고, 생식소가 복부와 위를 압박하여 먹이섭취가 어려워지므로 산란기 동안 섭식을 줄이거나 멈추어 산란준비에만 집중하는 것으로 알려져 있는데(Bond, 1979; Balanov et al., 2006), 벌레문치의 경우 다른 어류들에 비해 복강에서 생식소가 차지하는 비중이 높은 어종으로 산란시기인 11-2월이 되면서 생식소가 크게 증가하면서 다른 내장기관과 위를 압박하게 되어 다른 시기에 비해 공위율이 증가하게 되며, 먹이를 섭식하는 개체들도 많은 개체를 섭식하기 보다는 먹갈치와 청자 갈치처럼 크기가 큰 어류를 소량만 선택적으로 섭이하는 것으로 판단된다.