The characteristics of the oceanographic environment in the Aleutian Basin of the Bering Sea during spring in 1996, 1997, and 1999 were clarified. An investigation of the water properties revealed five basic layers in the Bering Sea during spring: (1) a surface layer of warm and low-salinity water induced by solar heating, (2) a subsurface layer of cold and low-salinity water propagated slowly by heat from the surface layer, (3) a thermocline layer where salinity was constant but temperature sharply decreased, (4) a temperature inversion layer, and (5) a deep layer with a gradual decrease in temperature and increase in salinity toward the bottom. The ranges of water temperature and salinity were 1.8-5.5℃ and 31.81-34.08 in 1996, 1.5-7.2℃ and 31.9-34.06 in 1997, and 0.5-5.6℃ and 32.0-34.11 in 1999, respectively. The water temperature of the surface layer was approximately 1.6℃ higher in 1997 than in 1996 and 1999. The lowest temperature at a depth of 100-150 m was about 1℃ lower in 1999 than in 1996 and 1997. Nutrient levels (nitrate, phosphate, and silicate) contributing to the control of the growth of phytoplankton were higher in the Aleutian Basin than in the eastern continental shelf and Bogoslof Island area. This was closely associated with the phytoplankton distribution. Nutrient concentrations were lowest at a depth of 25 m. The high primary production at that depth was confirmed from the vertical distribution of chlorophyll a. Chlorophyll a levels were above 4.0 μL-1 in some areas in 1996 and 1999, but below 2.0 μL-1 in most areas in 1997. Zooplankton density was about three times higher in 1999 than in 1997.
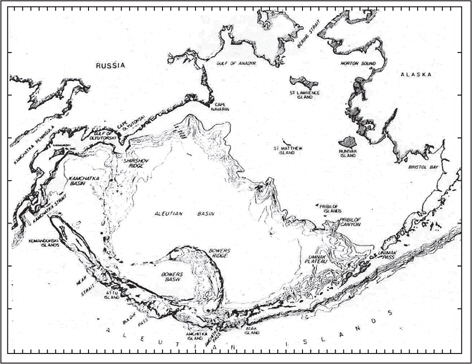
베링해는 Fig. 1과 같이 태평양의 북쪽에 위치하고 있으며 (Sayles et al., 1979), 동쪽의 알라스카(Alaska)반도, 서쪽의 캄차카(Kamchatka)반도, 남쪽의 알류산열도(Aleutian Islands) 그리고 북쪽의 베링해협(Bering strait) 등으로 둘러싸인 면적 2.3×106 km2, 체적 3.7×106 km3의 광활한 아한대 해역이다(Roden, 1995). 또한 베링해에는 수심이 3,000 m 이상인 거대한 알류산 해분(Aleutian Basin)이 중서부에, 보월스 해분(Bower Basin)과 캄차카 해분(Kamchatka Basin)이 서부에 형성되어 있으며, 동북∙부에는 수심 200 m 이하의 대륙붕이 베링해협을 포함하여 북쪽의 척치해(Chukchi Sea)까지 넓게 발달되어 있다(Kinder et al., 1980). 해수교환은 캄차카 해협(Kamchatka Strait), 니어 해협(Near Strait) 및 크고 작은 알류산 수로(Aleutian Pass)를 통하여 주로 이루어지고 있으며, 알류산 수로 중에서 암치트카 수로(Amchitka pass)는 1,000 m보다 얕은 상층 수의 해수교환이, 캄차카 해협은 1,000-2,000 m의 중층 수해수교환이 이루어진다(Favorite, 1967; Favorite, 1974; Reed et al., 1993; Roden, 1995).
베링해 내의 흐름은 알라스카 남쪽 해역에서 알라스카 환류(Alaska Gyre)가 발달되어 생성된 알라스카 해류(Alaska Stream)가 알류산열도의 남쪽 면을 따라 서쪽으로 흘러 니어 해협, 암치트카 수로 및 아묵타 수로(Amukta pass)를 통하여 베링해로 유입되며(Reed et al., 1993), 이는 베링해에 주요한 상부 대양 순환수이며, 알라스카 대륙으로부터 유입된 담수로 인하여 고온∙저염의 특징을 가지고 있음으로서, 이것의 세기가 베링해 순환에 영향을 준다고 하였다(Reed and Stabeno, 1994). Reed et al. (1994)의 결과에 의하면 1993년 4-5월의 관측 결과로부터 계산된 geopotential 지형의 양상이 동부 해역 200-400 m에 분포하는 4℃ 이상의 고온수가 알라스카 해류로부터 아묵타 수로를 통하여 공급됨을 나타내었으며 이러한 고온수는 명태의 산란 장소나 해수의 부력 조건에 의한 난의 수직 이동 또는 사망률 등에 영향을 주기 때문에 생태계의 조정이란 점에서 중요한 물리현상이라고 하였다.
베링해 내의 해류는 알류산열도의 북쪽을 따라 동쪽으로 흐르는 해류, 동부 베링해 대륙사면을 따라 북서쪽으로 흐르는 베링사면류(Bering Slope Current) 및 베링사면류로부터 기원이 됨을 확인한 캄차카 해류(Kamchatka current) (Verkhunov and Thachenko, 1992)가 존재한다. 또한 적은 규모의 환류나 와동(eddy)들이 대륙사면을 따라 형성되며(Coachman, 1986; Kinder et al., 1986; Reed et al., 1993), 대륙붕과 대륙사면이 만나는 지점에 염분의 차이에 의한 염분 전선이 형성되는데, 이는 강물의 유입이나 얼음이 녹아서 형성된 저염의 연안수(31.0-32.7)와 외해로부터의 고염 해수>32.7)가 혼합하면서 일어나는 현상이다. 이와 같은 물리적 기작에 의해 대륙사면을 따라 계속적으로 해수의 용승(upwelling)과 혼합(mixing)이 이루어져서 와류의 바깥쪽에 계속 영양염이 공급되고, 염분이 낮아져서 표층수괴의 안정으로 인해 식물플랑크톤이 성장하기 좋은 조건이 형성된다(Hansell et al., 1989). 조류는 대륙붕과 알류산 수로 등에서 강하게 나타나 4 m/sec가 되기도 하며, 바람은 에너지가 크고 변화가 심하여 표층 전선(surface fronts), 소용돌이(eddies)를 형성하고 국지적인 용승(upwelling) 및 침강(downwelling)을 야기 시킨다.
이 해역은 북극권에 위치함으로서 겨울이 되면 해양이 결빙되고 3-4월에 최대로 확장하였다가 봄이 되면 얼음이 서서히 녹고 여름에는 완전히 소멸되는 데 이러한 해빙의 생성과 소멸이 이 해역의 해양생태계에 커다란 영향을 미친다(Niebauer and Alexander, 1985). 또한, Parkinson et al. (1986)에 의하면, 베링해의 해빙은 겨울철에는 대륙붕까지 확장되었다가 4월경 서서히 녹기 시작하여 6월말에서 9월말까지 베링해협을 거쳐 북극해까지 녹아 베링해 표층 해수의 수괴안정도에 영향을 끼친다. 이 수괴의 안정도는 식물플랑크톤의 성장에 영향을 끼쳐 춘계 및 초여름에 blooming이 이루어지며, 이로 인한 영양염의 고갈과 외양 및 저서층으로의 탄소공급이 증가되어 동물플랑크톤 및 저서생물들의 생산력에 영향을 주게 된다고 하였다. 이러한 현상을 Grebmeier et al. (1995)는 베링해 생태계의 개념적 모델로서 표현하였다. 이 모델에 의하면 베링해에서의 식물플랑크톤 생산력은 계절적으로 변화하는 빛 조건 및 해빙의 위치와 밀접하게 연관되어 있으며, 식물플랑크톤의 생산력에 따라 종속영양 생물들 즉 박테리아, 원생동물 및 동물플랑크톤이 영향을 받는다고 하였다. 또한, 계절적으로 변화하는 해빙의 분포에 의하여 베링해의 해양학적 환경이 변동되므로 해빙의 분포가 베링해의 일차생산력을 좌우하는 주요 요인이라고 하였다. 따라서 동 해역은 계절별로 해양환경의 변동 및 생물의 분포형태가 계절별로 뚜렷이 나타나는 해역이며, 동계에는 해양의 결빙이 일어나고, 춘계 및 하계에는 인근의 육지나 북태평양으로부터 영양염류의 공급을 받고 기후조건도 양호함으로 생물체들이 번성한다. Walsh and McRoy (1986)는 베링해가 세계에서 가장 비옥한 해양 중의 하나이며, 특히 남동 대륙붕은 생물체들이 번성하여 대부분의 수산 어종들이 먹이에 제한을 받지 않고 성장∙번식할 수 있는 해역이라고 하였다.
베링해에서 생태계를 좌우할 수 있을 정도로 아주 풍부하고 중요한 어종은 회유성이 강하고 냉수성 어류인 명태(
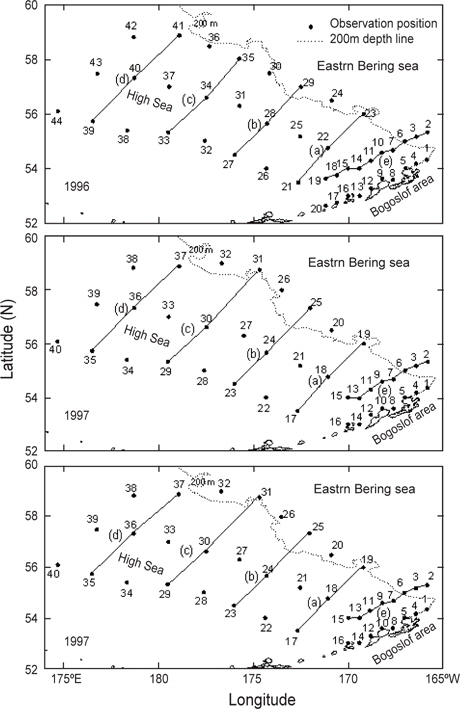
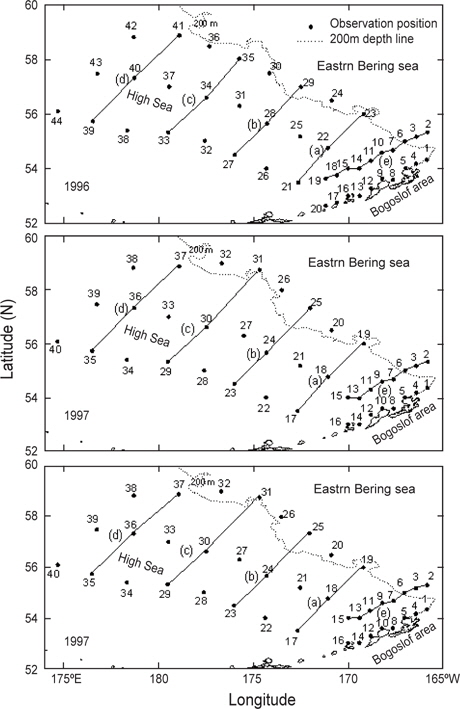
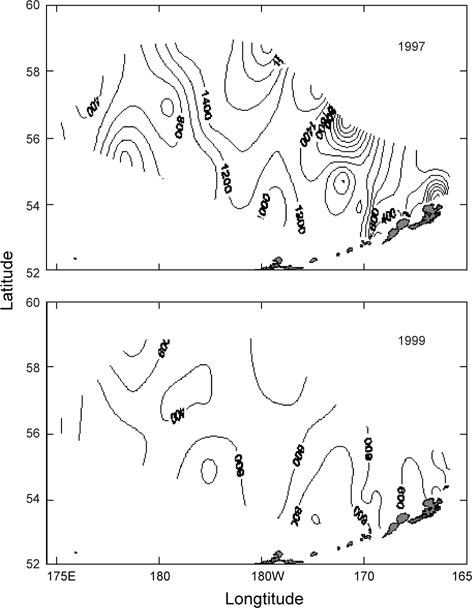
조사는 1996년 5월 10일부터 5월 31일까지, 1997년 5월 15일부터 6월 12일까지, 1999년 5월 15일부터 6월 17일까지 3개년에 걸쳐 베링해의 보고슬로프해역과 알류산 해분 및 동부대륙붕 해역의 해양학적, 화학적 및 생물학적 해양환경을 구명하기 위하여 국립수산과학원(구, 국립수산진흥원) 시험조사선 부산851호(1996년 및 1997년) 및 탐구1호(1999)를 이용하여 조사를 실시하였다(Fig. 2). 해양환경을 관측한 위치는 1997년과 1999년은 동일하였으나 1996은 약간의 차이가 있었다. 관측 정점은 1996년에 44개, 1997년 및 1999년에 40개였으며, 해역별로 보면, 1996년은 보고슬로프해역 20개, 대륙붕해역 2개, 베링공해 및 중간해역 22개였고, 1997년 및 1999년에는 보고슬로프해역 16개, 대륙붕해역 6개, 베링공해 및 중간해역 18개였다. 보고슬로프해역에서 연안의 수심은 200 m이천이고, 외해의 수심은 1500-2000 m 이며, 대륙붕해역은 수심 150 m이천이고, 베링공해 및 중간해역은 수심 3000-3500 m 이었다.
춘계 베링해의 물리학적, 화학적 및 생물학적 특성을 분석하기 위하여, 관측정점에서 수온 및 염분은 3개년간 CTD (Sea Bird Inc.)를 이용하여 표층에서부터 수심 500 m까지 수심 1 m 단위로 조사하였으며, 투명도는 1997년 및 1999년에 하얀색 원판인 Secchi disk (지름 35 cm)를 이용하여 주간에 실시한 관측 정점에서만 조사하였다.
영양염(용존무기질소, 인산인 및 규산규소)은 생물의 기초생산에 영향을 주는 중요한 인자이다. 따라서 춘계 베링해에 영양염의 정도를 파악하기 위하여 1999년에 모든 관측정점에서 6개의 수심층(0, 25, 75, 100, 200, 400 m)에 대해 2.5 L의 Losette sampler를 이용하여 해수를 채수하였으며, 간혹 표층해수는 용기를 이용하였다. 해수에 포함된 식물 및 동물플랑크톤을 제거하기 위하여 200 μm, 0.45 μm의 Waterman-GF/F 필터로 여과한 해수 500 mL를 용기에 담아 -30 ℃에 냉동 보관하였다가 실험실에서 Technicon AA-Ⅱ 자동분석기를 사용하여 분석하였다(Parsons et al., 1984). 용존무기질소(Dissolved inorganic acid)는 질산질소와 아질산질소를 합하여 분석하였다.
클로로필 a (㎍L-1)는 식물플랑크톤의 양을 정량적으로 표현하기 위해 주로 사용되는 지표이다. 이를 분석하므로서 춘계 베링해의 기초생산력 정도를 파악할 수 있다. 1996년 및 1997년에는 용기를 이용하여 표층해수만을 채수하였고, 수심별 클로로필 a의 변화를 파악하기 위하여 1999년에는 표층-400 m까지 2.5 L용 Losette sampler를 이용하여 6개 수심층(표층, 25, 75, 100, 200, 400 m)의 해수를 채수하였다. 채수한 시료는 즉시 0.45 μm Whatman-GF/F 필터를 사용하여 여과하여 알루미늄 호일에 싸서 냉동 보관하였다. 시료는 연구실에서 90% 아세톤으로 추출하고 형광분광광도계(Hitachi F-2000)에 의하여 형광법으로 분석하였다(Parsons et al., 1984).
동물플랑크톤은 해양관측정점과 동일한 지점에서 망 입구직경 60 cm, 바스켓 연결부 직경 11 cm, 망목 335 μm인 봉고네트를 사용하여 채집하였다. 채집방법은 표층에서 수심 100 m까지 약 3-3.5 노트로 예망하면서 경사채집 하였으며, 예망시간은 약 5분 정도로 하였고, 정량분석을 위하여 네트 입구에 유속계(Digital flow meter)를 부착하였다. 채집된 표본은 선상에서 중성 포르말린 (5-10%)으로 고정하였다. 고정된 표본은 실험실에서 수분을 빼고 전체중량을 측정한 다음, 예망시 얻어진 여과량을 적용하여 m3당 중량으로 환산하였다.
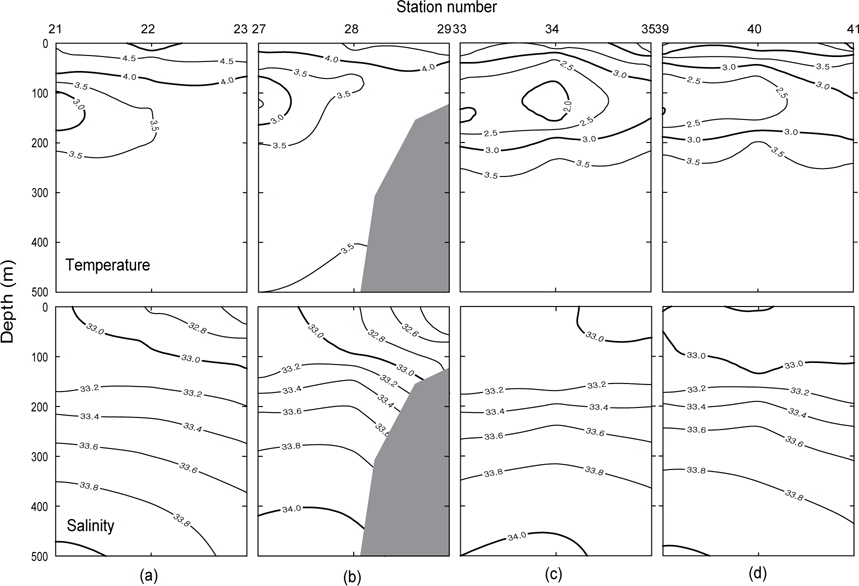
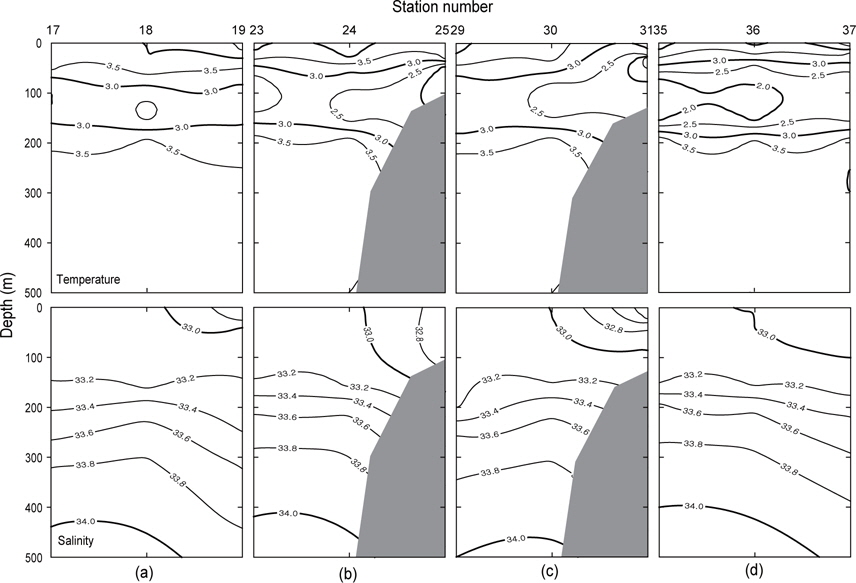
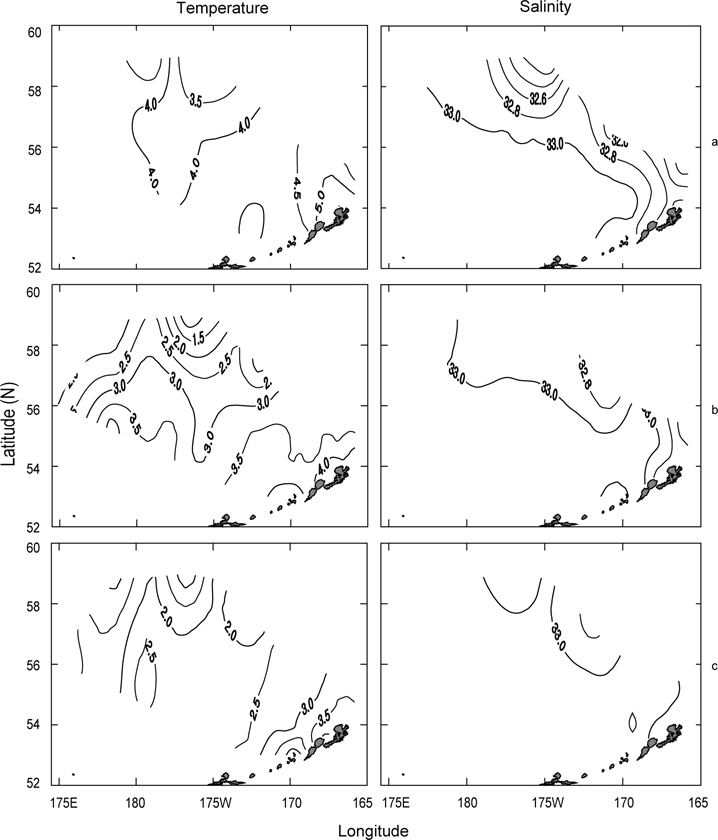
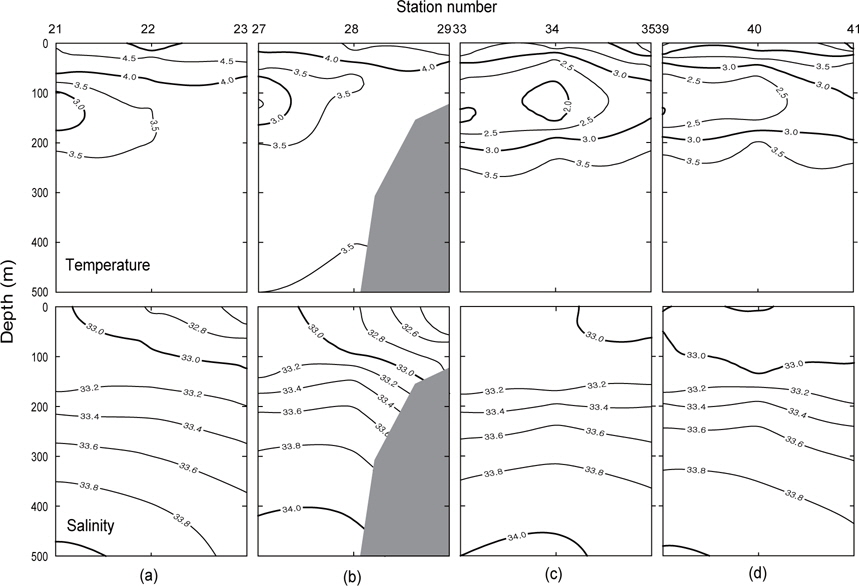
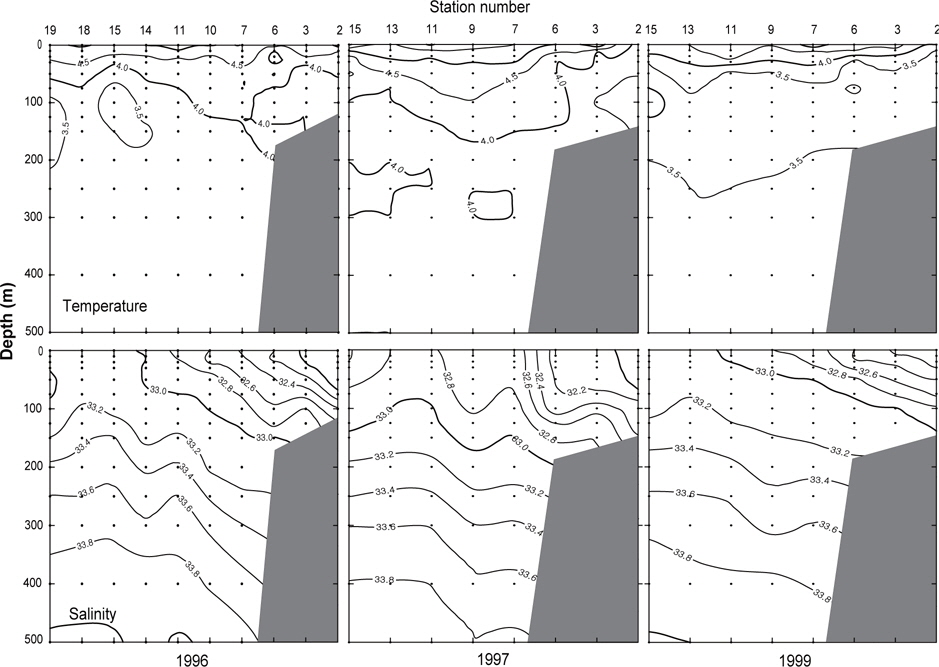
Fig. 3은 1996년 조사해역 (Fig. 2)에서 단면 a, b, c, d에 대한 각각의 연직 수온 및 염분을 나타내었다. 각 단면에서의 연직 수온은 표층에서 수심 75 m까지 수심이 증가함에 따라 수온이 낮아지는 패턴을 보이고, 수심 75 m에서 150 m사이에서는 core형으로 나타나며, 동쪽이 높고 서쪽이 낮은 동고서저형이며, 최소수온이 나타나는 수층이다. 그리고 수심 150 m에서 수심 250 m까지는 수심이 증가함에 따라 수온이 증가하는 패턴을 보이며, 수심 250 m이심에서는 3.5℃로서 수온의 변동이 거의 없다. 이러한 현상은 하계에 행해진 조사결과에서도 같은 결과를 나타내고 있다(Kim, 1998). 이는 베링해에서의 계절적 변동에 따른 연직 수온의 변동은 표층에서 수심 250 m까지에서 주로 발생하고 있음을 알 수 있다. 또한 단면 a에서 단면 d로 가면서 수심 75-150 m 사이의 수온이 낮아지는 패턴을 보이고 있다. 연직 염분은 대체적으로 수심이 증가함에 따라 염분이 증가하는 패턴을 보이고 있다. 단면 a (정점 23) 및 b (정점 29)에서는 염분이 대륙붕에서 외해쪽으로, 표층에서 수심 아래로 확산되는 현상이 뚜렷하게 나타나고 있으며, 표층의 염분은 동쪽에서 32.8이하로 나타났고, 서쪽에서는 33.0 이상의 염분을 나타냈다. 그러나, 단면 c 및 d에서는 단면 a 및 b에서 나타나는 현상이 거의 나타나지 않고 있다. 이것은 단면 c에서의 정점 35와 단면 d에서의 정점 41이 대륙붕에서 떨어져 외해역에 위치하고 있기 때문인 것으로 판단된다.
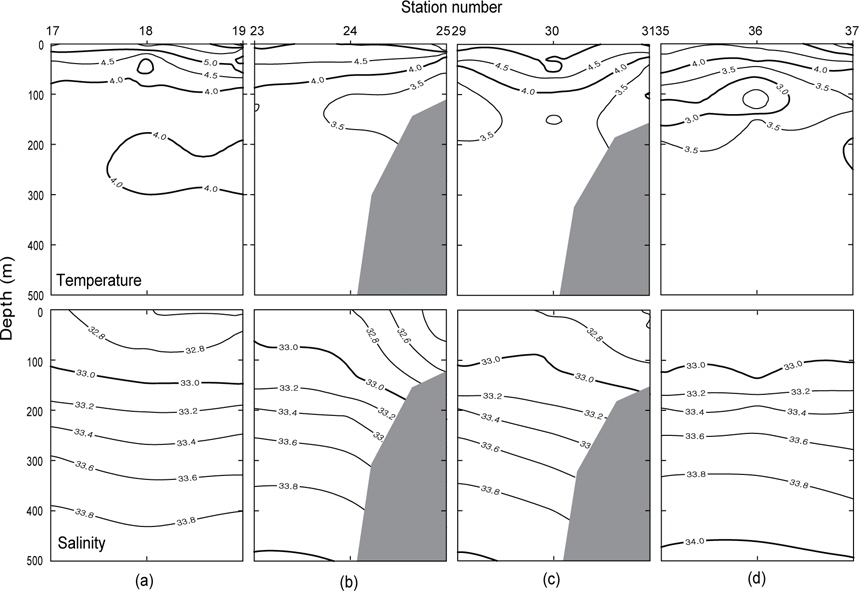
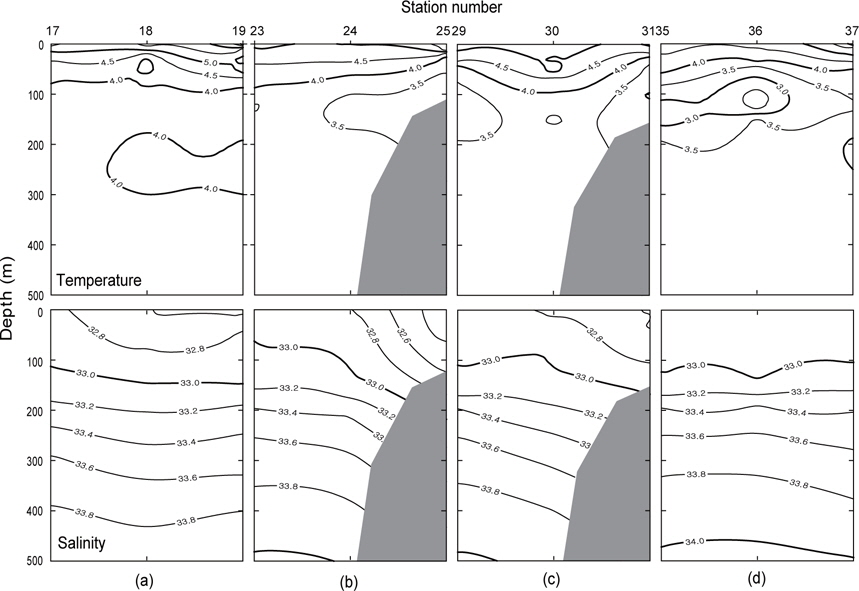
1997년 조사해역 Fig. 2에서 단면 a, b, c, d에 대한 각각의 연직 수온 및 염분을 Fig. 4에 나타내었다. 1997년의 패턴은 1996년과 거의 동일하나, 수온은 1996년보다 0.5-1℃ 정도 높게 나타나고 있으며, 염분은 약간 낮은 농도를 나타내고 있다. 특히 3.0℃이하의 수온이 단면 d에서만 나타날 뿐 다른 단면에서는 전혀 나타나지 않고 있으며, 단면 a에서 수심 300-400 m사이에 4.0℃의 등온선이 뚜렷하게 보이고 있다. 1997년도 1996년과 마찬가지로 수심 100-150 m에서 최소수온을 나타내며, 단면 a에서 단면 d로 가면서 수온이 낮아지는 패턴을 나타내고 있다. 염분의 경우, 수심이 증가함에 따라 증가하는 패턴을 보이며, 1997년이 1996년보다 낮게 나타났다. 이는 단면 a에서 나타나는 현상을 비교해보면 보다 뚜렷하게 볼 수 있다.
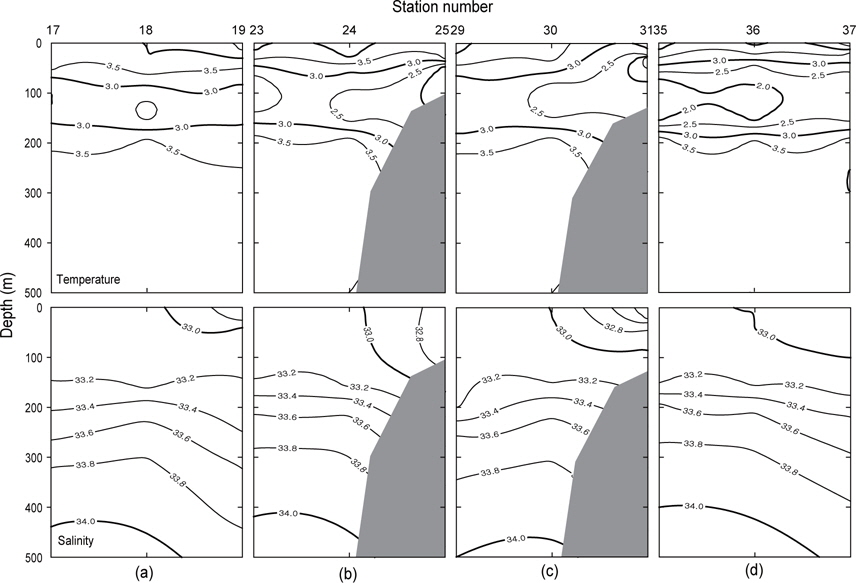
Fig. 5는 1999년 조사해역 (Fig. 2)에서 단면 a, b, c, d에 대한 연직 수온 및 염분을 나타내고 있다. 연직 수온은 표층에서 수심 150 m까지 1996년에 비하여 0.5-1.0℃ 낮게 나타나고, 1997년에 비하여 1.0-1.5℃ 낮게 나타났다. 특히 단면 b 및 c에서 2.5℃ 이하의 찬물이 대륙붕에서 외해로 뻗치고 있는 현상을 뚜렷하게 보여주고 있다. 단면 d에서는 2.0℃이하의 찬물이 외해역에서 대륙붕쪽으로 강하게 뻗치고 있다. 이로서 1999년에는 찬물이 대륙붕과 외해역에서 외해역의 중앙부로 뻗치고 있음을 알 수 있다. 앞에서 언급한 2개년과 마찬가지로 단면 a에서 단면 d로 이동함에 따라 수온이 낮아지는 패턴을 보이고 있으나, 최소 수온이 나타나는 수심 층에서는 두 해에 비하여 찬물이 넓게 분포하고 있다. 연직 염분은 1999년이 1996년 및 1997년에 비하여 0.2 정도 높은 농도를 보이고 있으며, 패턴은 수심이 증가함에 따라 높아지는 양상을 나타내고 있다. 특히 33.0의 등밀도선이 대륙붕 가까이에서 나타나는 것을 볼 수 있다.
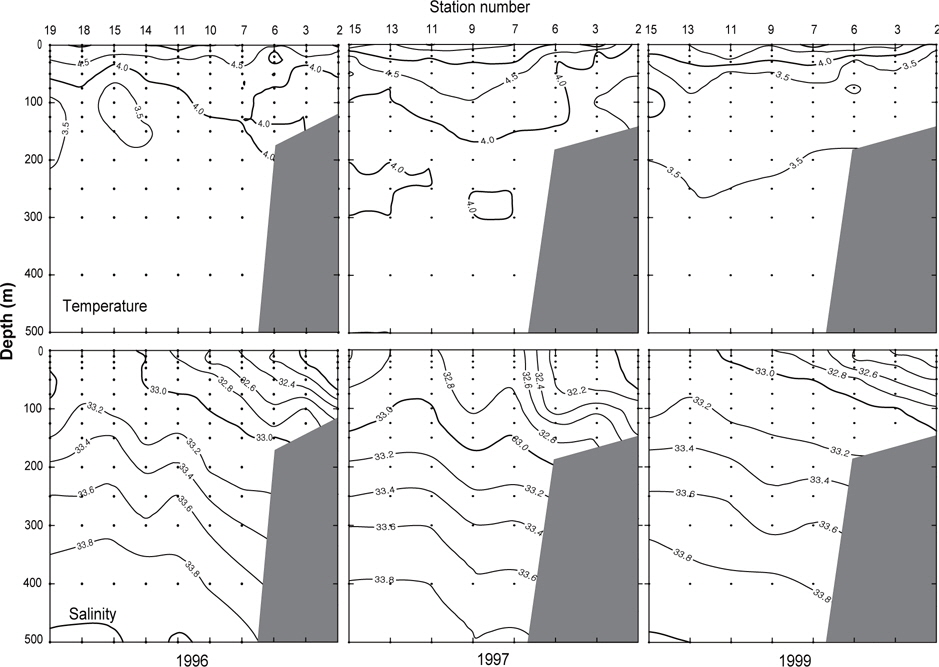
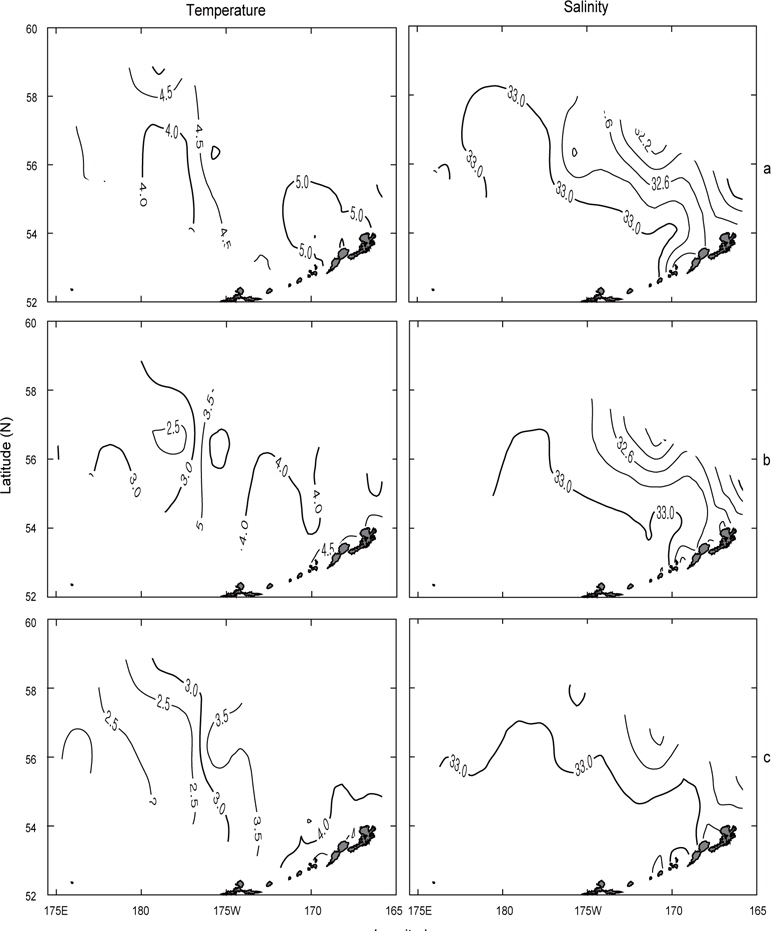
또한, 알류산 열도 근처 보고슬로프해역의 연직수온 및 염분의 분포를 알아보기 위하여 알류산 열도와 평행하게 단면을 이루고 있는 Fig. 2의 단면 e에 대한 연도별 수온 및 염분의 연직분포를 Fig. 6에 나타내었다. 단면 e에서의 수온은 앞에서 언급한 단면 a, b, c, d에서의 수온보다 높으며, 수온의 급격한 변화도 거의 없거나 아주 약하게 나타나고 있다. 표층수온은 동쪽이 낮고 서쪽이 높게 나타나고, 최소 수온은 1996년에 정점 14, 15 및 19의 수심 75-200 m사이에서 3.5℃이하로, 1997년에 정점 2의 수심 50-125 m사이에서 3.5℃이하로, 그리고 1999년에 정점 15의 수심 75-125 m사이에서 3.0℃이하로 나타났다. 염분은 동부대륙붕에 위치한 정점 2, 3 및 6에서 32.4 이하의 낮은 농도를 나타내고 있으며, 이것이 보고슬로프해역의 외해역으로, 또한 표층 아래로 확산되어 외해역의 중앙부까지 및 동부대륙붕의 저층까지 영향을 미치고 있다.
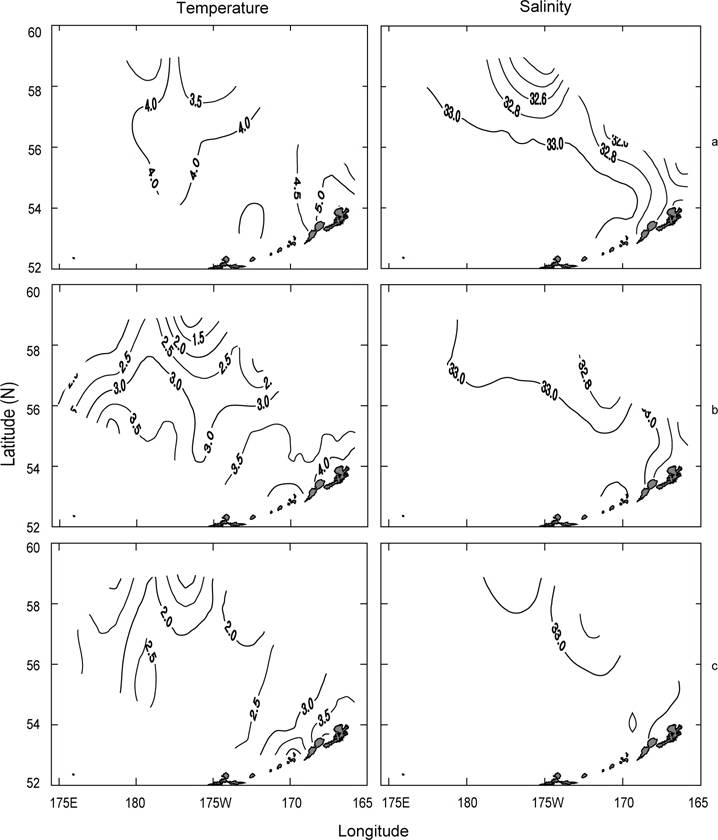
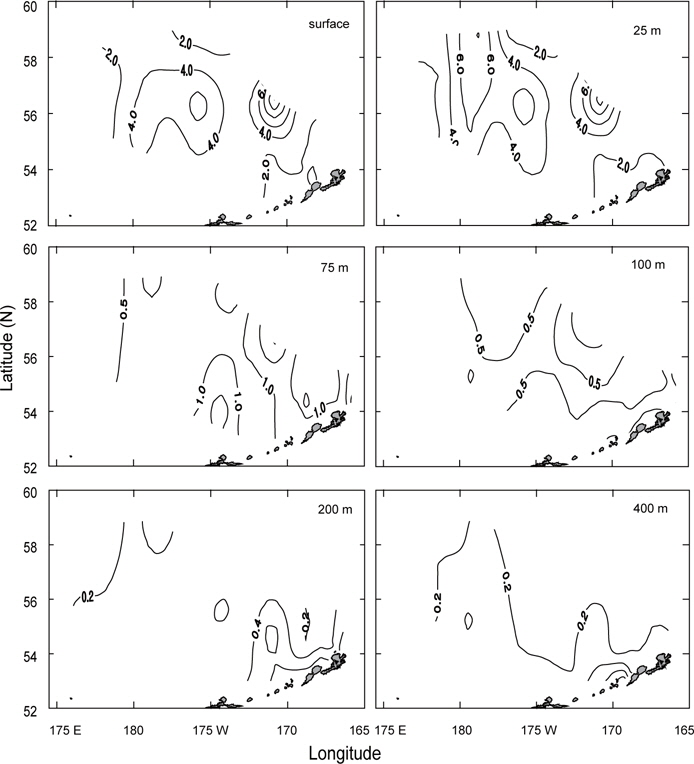
수온 및 염분의 수평분포는 표층, 수온이 급격히 낮아지는 50 m 수심층 및 최소 수온층이 나타나는 100 m 수심층에 대하여 연도별로 나타내었다. 연도별 분포패턴은 수심에 따라 다른 양상을 나타내고 있다.
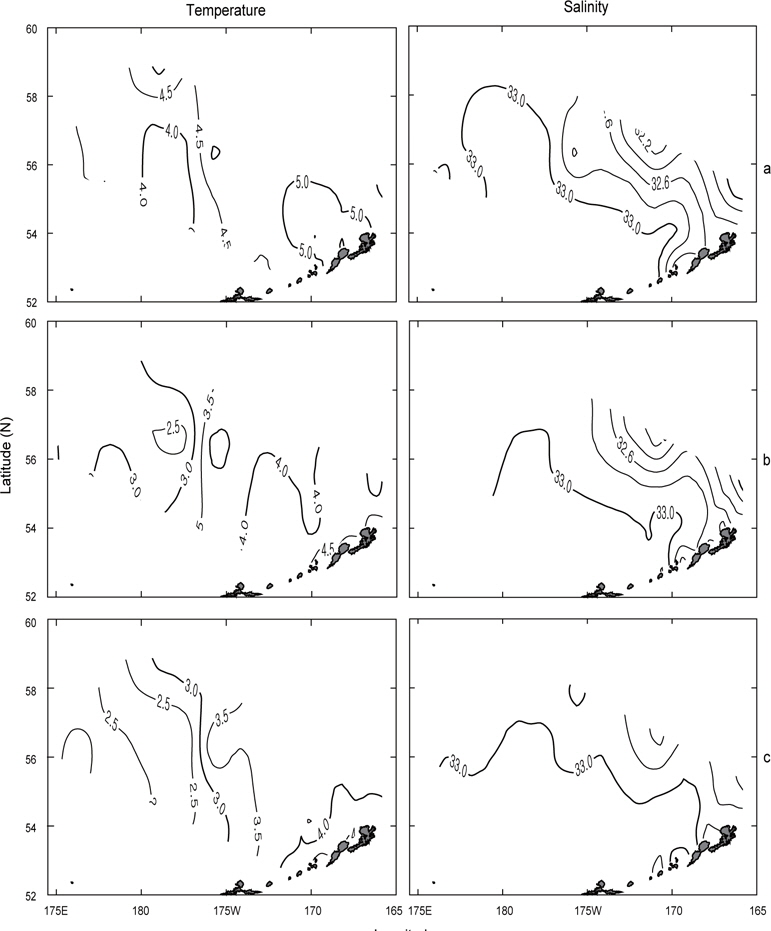
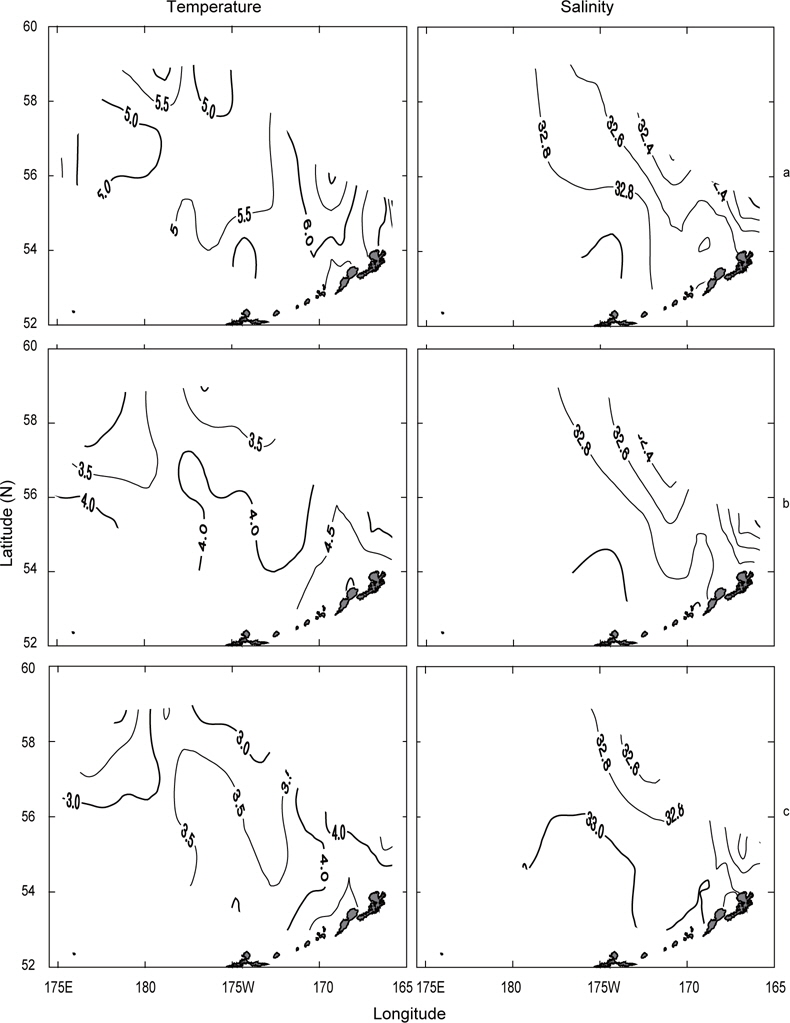
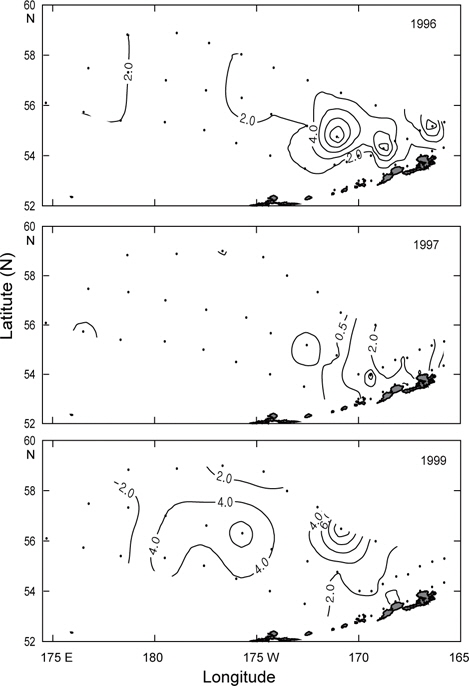
먼저 1996년의 수평분포를 Fig. 7에 나타내었다. 표층수온은 3.7℃ (정점 32)-5.5℃ (정점 5)의 범위였고, 5.0℃ 이상의 영역은 보고슬로프 해역 및 북쪽 끝에서, 4.0℃이하의 영역은 베링공해 중앙에서 각각 나타났다. 수온전선은 베링공해 중앙에서 약하게 보이고 있다. 표층염분은 북동쪽 끝에서 32.0 이하로 아주 낮게 나타났고, 분포패턴은 대륙붕에서 32.4 이하로 낮게, 외해에서는 32.8 이상으로 높게 나타났으며 염분전선이 대륙붕과 평행하게 형성되어 있다. 이것은 동계에 결빙되었던 저염의 대륙붕 해수가 녹으면서 일어나는 현상이다. 수심 50 m의 수온은 서쪽에서 예리하게 낮아져서 동쪽이 높고 서쪽이 낮은 동고서저의 형태를 띠고 있으며, 외해 중앙에 수온전선이 강하게 형성되어 있다. 보고슬로프 연안을 따라 나타나는 고온현상은 알류산 수로를 통하여 알류산 해류의 고온 해수가 유입되어 나타나는 현상이라고 판단된다. 수심 50 m의 염분은 표층과 거의 동일한 양상으로 나타나고 있으며, 수온과 마찬가지로 알류산 열도 근처에 알류산 수로를 통하여 고염이 유입되고 있음을 보여준다. 이는 Reed and Stabeno (1994)의 연구결과와 거의 일치하고 있음을 알 수 있다. 최소 수온층이 나타난 수심 100 m의 수온은 보고슬로프 연안을 제외하고 4.0℃ 이하로, 염분은 32.6 이상을 보여주고 있다. 수온전선은 여전히 외해 중앙에 있고, 염분의 분포로부터 대륙붕의 해수가 수심 100 m까지 영향을 주고 있음을 알 수 있다. 또한 수심 50 m에서 나타난 것과 마찬가지로 알류산 열도의 알류산 수로를 통하여 고온고염의 해수가 유입되고 있음을 볼 수 있다.
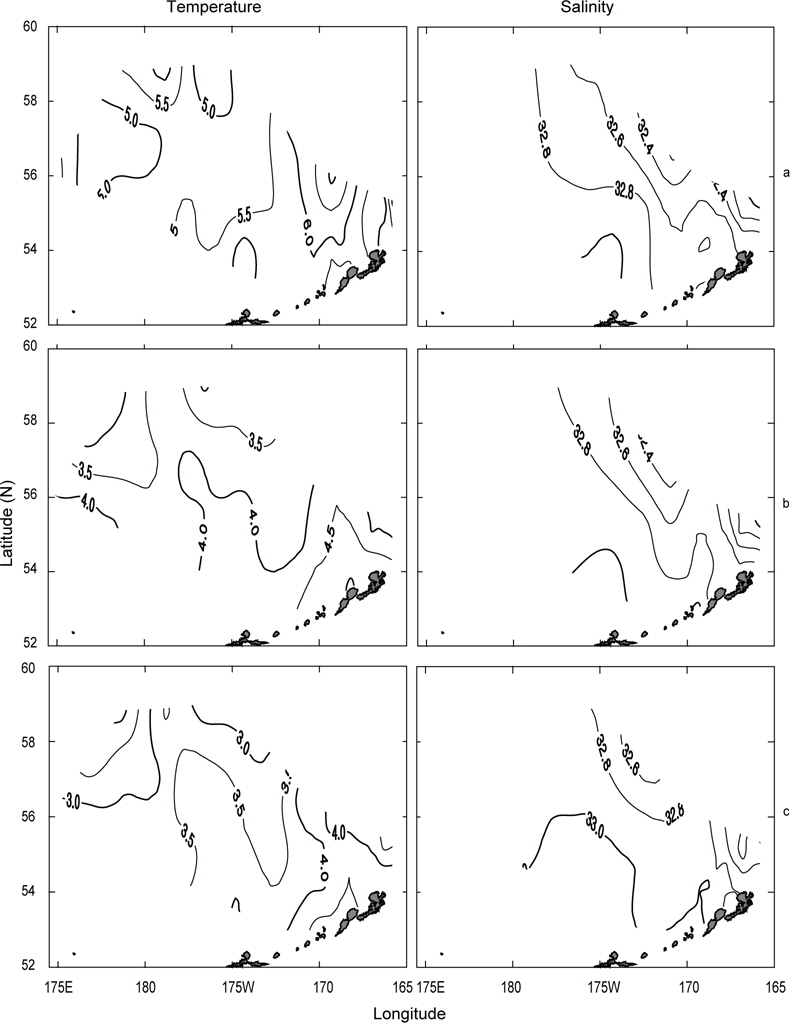
Fig. 8은 1997년의 수심별 수온 및 염분의 분포를 나타내었다. 표층수온 및 염분은 3.8℃ (정점 2)-7.2℃ (정점 19)의 범위로, 31.9 (정점 2)-33.07 (정점 22)의 범위로 각각 나타나, 1996년보다 수온은 평균 0.5℃이 높고, 염분은 조금 낮게 나타났다. 표층수온은 동쪽 끝 부분을 제외하고 5.0℃ 이상을 나타내며, 1996년도와 달리 동쪽 연안에서 알류산 열도의 직각방향으로 전선이 형성되어 있다. 표층염분은 33.0 이상의 영역이 외해역 남쪽에 아주 좁게 나타나고 있는 것 외에는 1996년의 양상과 거의 동일하였다. 수심 50 m의 수온은 보고슬로프해역에서 4.5℃이상으로, 베링공해 북서쪽에서 3.0℃ 이하로 나타나며, 북서에서 남동으로 갈수록 수온이 점점 증가하는 현상을 보이고 있다. 수심 50 m의 염분은 외해역에서 33.0 이상으로, 동부대륙붕 및 동쪽 끝에서 32.8 이하로 나타나며, 특히 약한 염분전선이 보고슬로프연안 직각으로 형성되어 있다. 이는 1996년과 같이 고온고염의 해수가 알류산 수로로 유입되고 있음을 보여주는 현상이다. 최소 수온층이 나타나는 수심 100 m의 수온은 수 심 50 m과 마찬가지로 남동쪽은 높고 북동쪽이 낮은 동고서저의 패턴을 보이고 있고, 보고슬로프 연안에서 4.5℃ 이상의 고온이 나타나고 있다. 염분은 패턴이 거의 동일하나 동부대륙붕에서 32.6 이하의 저염 현상이 나타나고 있다. 이 층에서도 알류산 수로를 통하여 고온고염의 해수가 베링해로 유입되는 현상을 보여주고 있다.
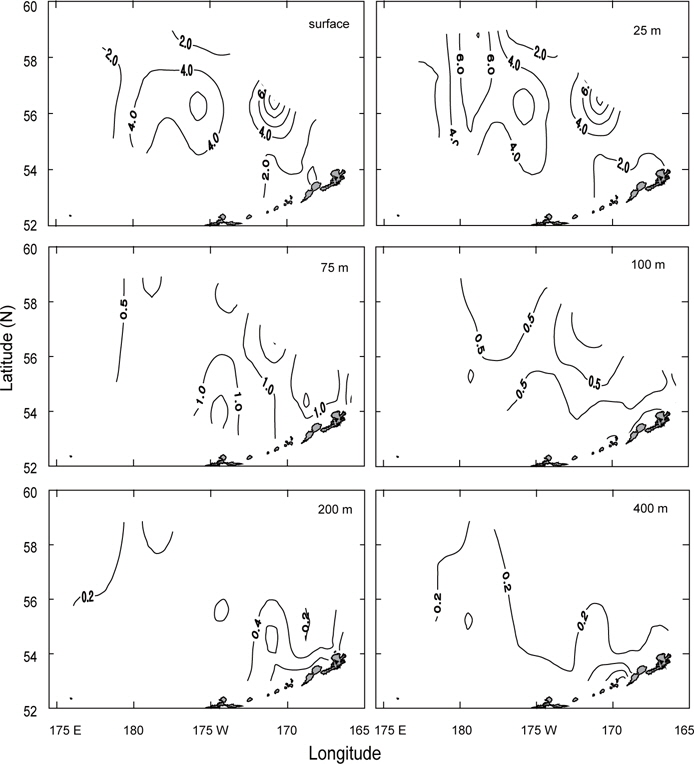
1999년의 수심별 수온 및 염분의 수평분포를 Fig. 9에 나타내었다. 표층수온과 염분은 3.0℃ (정점 32)-5.6℃ (정점 5)의 범위로, 32.13 (정점 2)-33.10 (정점 10)의 범위로 각각 나타났다. 평균수온은 1996년보다 0.4℃, 1997년보다 1.1℃ 낮았고, 평균염분은 1996년보다 0.1, 1997년보다 0.13 높았다. 1999년의 표층수온은 두 해와 달리 북쪽에 3.5℃ 이하의 저온수가 존재하며, 또한 염분도 32.6 이하의 저염수가 나타나고 있어 북쪽의 해수가 남쪽으로 흘러가는 것처럼 보인다. 수심 50 m의 수온은 베링공해 북쪽과 동쪽에 2.0℃ 이하의 저온수가 존재하며, 남쪽으로 내려갈수록, 동쪽으로 갈수록 점점 증가하는 경향을 나타내고 있다. 보고슬로프 연안은 4.0℃ 이상의 수온을 보이고 있다. 수온전선이 1996년 및 1997년보다 강하게 형성되고 있다. 염분은 전반적으로 32.8 이상의 고염을 나타내고 있으며, 특히 북태평양의 해수인 33.1의 고염수가 보고슬로프 연안의 알류산 수로를 통하여 유입되고 있는 현상을 뚜렷하게 보이고 있다. 최소의 수온층이 나타나는 수심 100 m의 수온은 베링공해에서 2.0℃ 이하의 등온선이 50 m보다 확장되어 나타나고, 보고슬로프해역에서는 3.0℃ 이상의 수온을 나타냈다. 염분은 대부분이 33.0 이상의 고염을 보이고, 특히 수심 50 m에서 나타났던 33.1의 등염분선이 대륙붕까지 강하게 뻗쳐있다. 이로서 수심 50 m 아래에서 보고슬로프 연안의 알류산 수로를 통하여 북태평양의 해수가 유입된다는 사실을 알 수 있다.
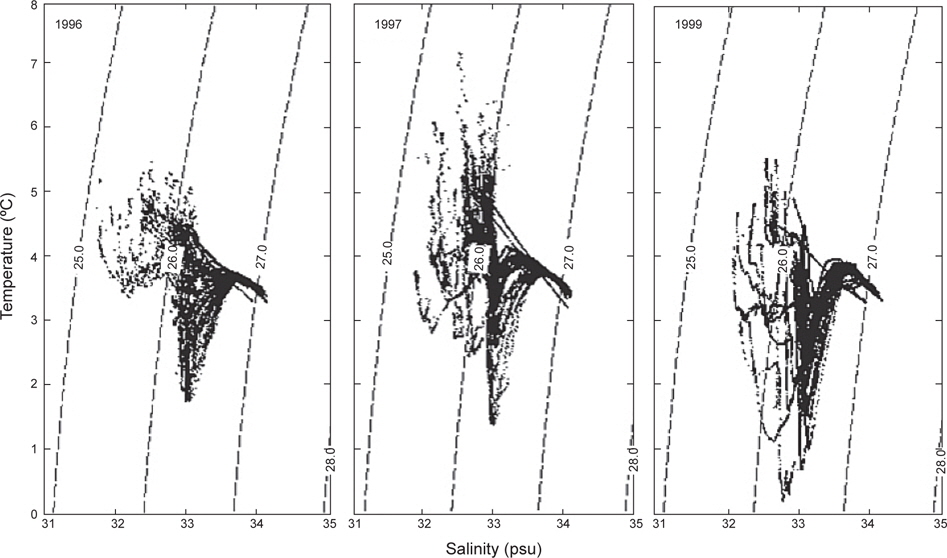
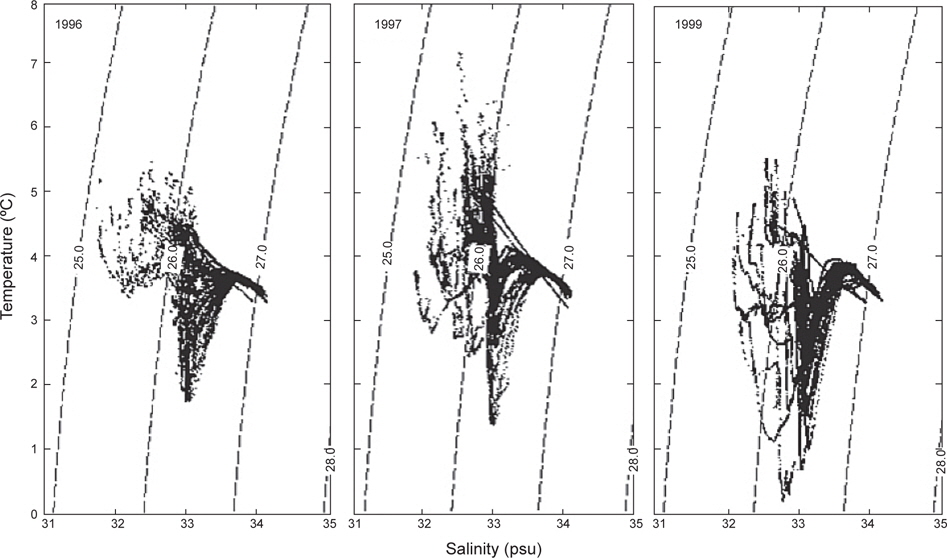
앞에서 수온 및 염분의 수평과 연직 분포를 살펴보았으며, 또한 3개년에 걸쳐 획득한 자료를 이용하여 춘계 알류산 해분의 수괴를 살펴보기 위하여 수온∙염분도(T-S diagram)를 Fig. 10에 나타내었다. 전체 패턴은 동일한 양상을 나타내고 있으며, 5가지의 수괴로 분류할 수 있다; 1) 태양열에 의해 수온이 상승하는 고온∙저염의 표층 수, 2) 표층으로부터 열의 전달이 천천히 이루어지고 있는 저온∙저염의 아표층 수, 3) 염분은 거의 일정하나 수온이 급격히 내려가는 수온약층, 4) 수온은 급격히 증가하나 염분이 조금씩 증가하는 수온역전층 및 5) 수온은 수심이 증가함에 따라 내려가나 염분은 점점 증가하는 수심이 깊은 층이다. 이러한 현상은 Sayles et al. (1979)의 관측결과와 거의 일치하고 있다. 정점별로 수괴를 살펴보면, 4가지의 그룹으로 나타낼 수 있다; 1그룹은 보고슬로프 연안에 위치한 정점으로 수온 약층이 거의 없거나 아주 약하게 나타나고 있으며, 2그룹은 보고슬로프 외해에 위치한 정점으로 1그룹보다 수온약층이 약간 강하게 나타나고, 3그룹은 동부대륙붕에 위치한 정점으로 33.0 이하의 저염의 수괴를 형성하고 있으며, 마지막 4그룹은 외해에 위치한 정점으로 수온약층이 아주 강하게 형성되어 있다.
수온과 염분의 범위는 1996년에 1.8-5.5℃, 31.81-34.08로, 1997년에 1.5-7.2℃, 31.9-34.06로, 1999년에 0.5-5.6℃, 32.00-34.11로 각각 나타났다. 1999년의 수온은 다른 두 해에 비하여 약 1℃ 이하로 낮게 나타났으나, 염분은 약간 높았다. 조사시기로 보았을 때 1999년이 1997년과는 거의 동일하게, 1996년보다는 약 15일 늦게 이루어졌음에도 불구하고 두 해에 비하여 낮은 수온을 나타내고 있다. 따라서, 베링해에서의 계절변동은 일정하지 않고, 연간변동이 일어나고 있음을 보여준다. 이는 동부베링해 대륙붕에서의 수온이 몇 도정도로 연간변동을 하고 있다는 보고와도 일치하고 있으며(Azumaya and Ohtani, 1995; Ohtani and Azumaya, 1995), 또한 이러한 현상이 1996년 및 1997년보다 1999년이 강하게 나타났음을 알 수 있다.
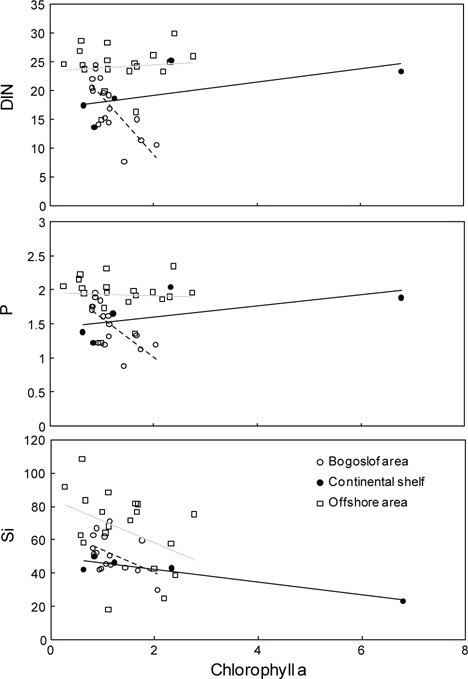
3년간의 걸쳐 분석한 표층 클로로필
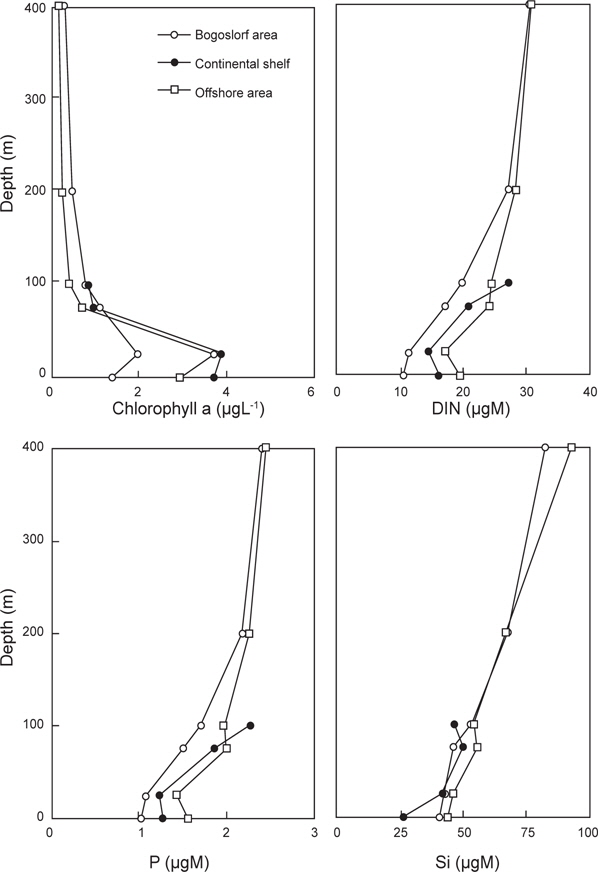
춘계 베링해에서 수심별 클로로필
클로로필
해역별 클로로필
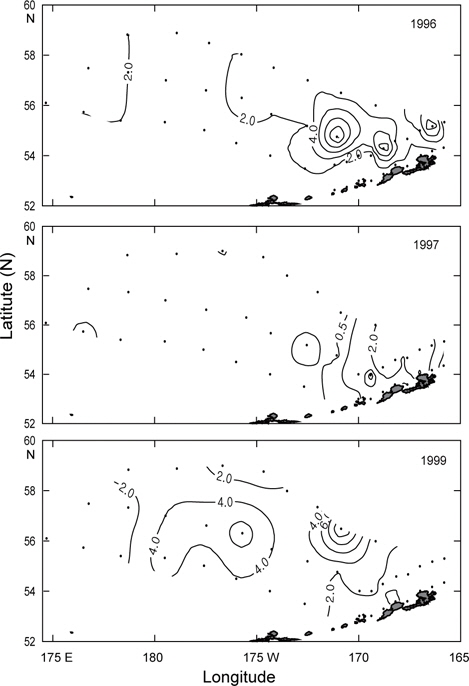
1997년과 1999년에 조사한 동물플랑크톤의 수평분포를 Fig. 15에 나타내었다. 1997년은 79 (정점 14)-3,167 mgm-3 (정점 20)의 범위를 나타내어 분포범위가 아주 넓고, 1999년은 250 (정점 4)-1,000 mgm-3 (정점 17)의 범위로 분포범위가 좁게 나타났다.
1997년의 동물플랑크톤은 베링공해 서쪽 끝과 보고슬로프해역을 포함한 동쪽 끝에서 600 mgm-3의 낮은 분포를 나타내고, 그 외 해역에서는 1,000 mgm-3이상의 높은 분포를 보이고 있다. 특히 보고슬로프 연안에서는 400 mgm-3이하의 아주 낮은 분포를 나타내었다. 1999년은 알류산 해분 해역 남부에서 800 mgm-3이상의 분포를 보이나, 베링공해 중앙에서는 400 mgm-3 이하의 낮은 분포를 나타내고 있다. 2개년의 분포패턴을 보면, 1997년은 대륙붕해역에서 알류산 해분 해역으로 이동하는 형태를 띠고 있고, 1999년은 알류산 해분 해역 남쪽에서 중앙으로 이동하는 형태를 띠고 있다. 또한 1997년의 동물플랑크톤 분포량은 1999년에 비하여 약 3배정도 높게 나타났으며, 이것은 계량어군탐지기에 나타난 표층에서부터 수심 50 m까지 밀집되어 나타났던 기록이 1997년이 1999년보다 훨씬 진하게 나타났던 것과 일치하고 있다. 동물플랑크톤의 분포는 클로로필
따라서 춘계 베링해에서의 동물플랑크톤의 분포는 거의 동일한 시기라 할지라도 해양환경(수온 및 염분)의 조건 및 클로로필
이상의 결과를 정리하여 보면, 1996년, 1997년 및 1999년 3년간에 걸쳐 국립수산과학원(구, 국립수산진흥원) 시험조사선 부산 851호 및 탐구 1호를 이용하여 자료를 수집하고, 수집한 자료를 분석하여 춘계 베링해 알류산 해분의 해양환경 특성을 구명하였다. 3년간의 조사결과로부터 동 해역의 수괴를 1) 태양열에 의해 수온이 상승하는 고온∙저염의 표층 수, 2) 표층으로 부터 열의 전달이 천천히 이루어지고 있는 저온∙저염의 아표층수, 3) 염분은 거의 일정하나 수온이 급격히 내려가는 수온약층, 4) 수온은 급격히 증가하나 염분이 조금씩 증가하는 수온 역전층, 5) 수온은 수심이 증가함에 따라 내려가나 염분은 점점 증가하는 수심이 깊은 층 등 5가지로 분류하였으며, 이것은 Sayles et al. (1979)의 관측결과와도 일치하고 있다. 3년간의 표∙중층에서 나타나는 복잡한 수괴와 수심 300 m 이하에서 나타나는 거의 동일한 수괴는 동계에 동부대륙붕 표층해수가 결빙되었다가 계절이 춘계로 변화면서 태양열에 의하여 얼음이 서서히 녹기 시작하여 저온∙저염수가 대륙붕에서 외해로 이동하면서 나타나는 것과 알류산열도를 따라 동쪽에서 서쪽으로 흐르는 고온∙저염의 북태평양 해수가 알류산 수로를 통하여 유입되면서 일어나는 현상이며, 이러한 원인의 강약에 따라서도 수괴의 패턴이 다르게 나타날 것으로 생각한다. 이것은 Stabeno and Reed (1994)에 의한 베링해에서의 평균순환 및 Overland et al.(1994)에 의한 수치적 buoy 추적 모델링을 이용한 표층 순환 연구의 결과로부터 유추 해석할 수 있다.
수온과 염분의 분포범위는 1996년에 1.8-5.5℃, 31.81-34.08로, 1997년에 1.5-7.2℃, 31.9-34.06로, 1999년에 0.5-5.6℃, 32.00-34.11로 각각 나타났다. 표층수온은 1997년이 1996년 및 1999년에 비하여 약 1.6℃ 높게 나타났고, 수심 100-150 m 사이에서 나타나는 최소수온은 1999년이 다른 두 해에 비하여 약 1℃ 이하로 낮았다. 조사시기로 보았을 때 1999년이 1997년과는 거의 동일하게, 1996년보다는 약 15일 늦게 이루어졌음에도 불구하고 두 해에 비하여 낮은 수온을 나타내고 있다. 이는 베링해에서 계절변동의 시기가 일정하지 않고, 연간변동이 일어나고 있음을 보여준다. 이는 동부베링해 대륙붕에서의 수온이 몇 도정도로 연간변동을 하고 있다는 보고와도 일치하고 있다(Azumaya and Ohtani, 1995; Ohtani and Azumaya, 1995). 표층염분은 동부대륙붕에서 낮고, 알류산 해분 해역에서 높은 분포를 나타내었고, 수심 300 m 이하의 층에서는 수심이 증가함에 따라 염분도 증가하는 3개년 공히 동일한 분포패턴을 보였다.
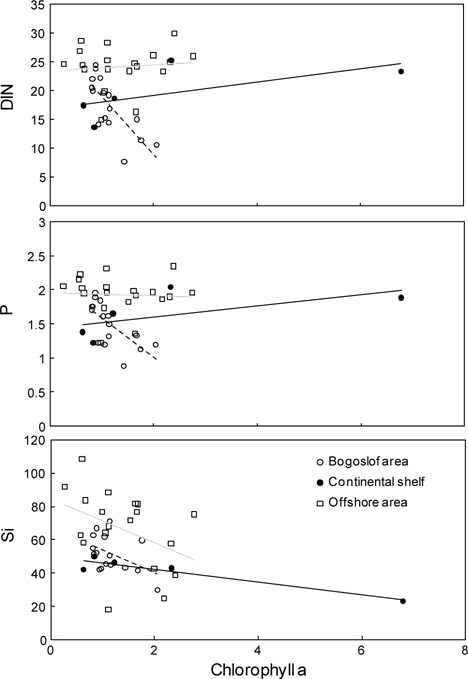
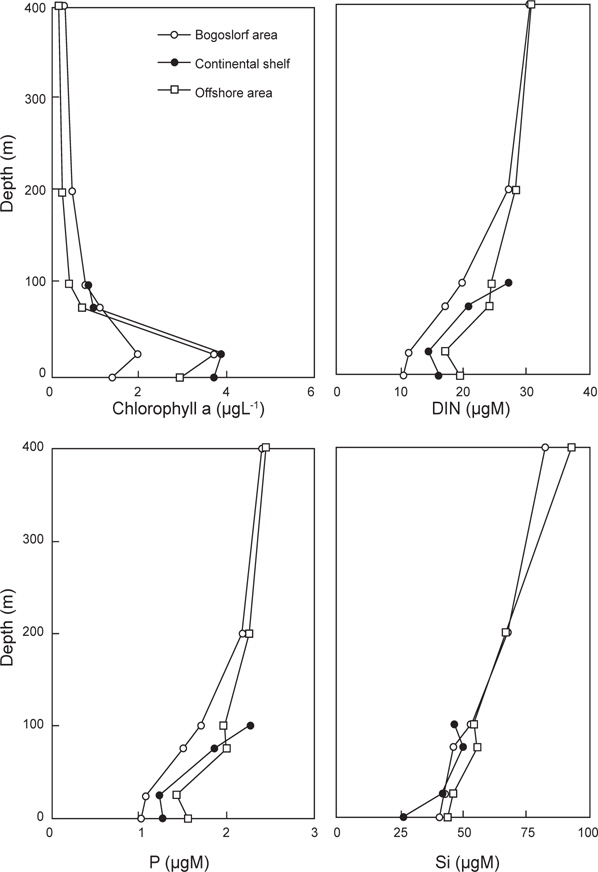
식물플랑크톤의 성장 조절에 관여하는 용존무기질소, 인산인 및 규산규소는 알류산 해분 해역에서 가장 높고, 동부대륙붕, 보고슬로프해역 순으로 나타났다. 또한 수심 25 m층에서 가장 낮은 분포를 보였다. 이것은 식물플랑크톤의 번성과 밀접한 관계가 있는 것으로 생각된다. 따라서 베링해에서 표층보다는 표층 20 m 아래층에서 기초생산력이 활발하게 이루어지고 있음을 알 수 있었다.
클로로필 a는 식물색소 중 양이 가장 많으며 활성 또한 강한 것으로 알려져 있으므로 기초생산량 변동에 이용되고 있다(Dring, 1992). 1996년 및 1999년에는 4.0 ㎍L-1 이상의 높은 분포를 나타낸 해역이 있는 반면에 1997년에는 2.0 ㎍L-1 이하의 낮은 분포를 나타내었다. 이는 식물플랑크톤을 주 먹이로 하는 이차생산자인 동물플랑크톤의 증식과 밀접한 관계가 있는 것으로 생각된다. 왜냐하면, 1997년과 1999년의 동물플랑크톤 양을 비교해 보면, 1997년이 1999년에 비하여 약 3배 높은 양을 나타내고 있기 때문이다. 또한 동∙식물플랑크톤이 번식할 수 있는 해양학적 조건과도 밀접한 관계가 있다고 생각된다.
이러한 결과는 해양생태계의 물질순환과 먹이구조에서 일차생산은 대부분이 식물플랑크톤에 의하여 이루어지며, 동물플랑크톤의 먹이인 유기물은 원천적으로 식물플랑크톤의 일차생산에서 기원한다는 설을 입증하고 있다. 또한, 베링해에서의 생물학적인 패턴은 전적으로 계절에 따라 변화하는 물리적, 생물학적 현상과 밀접하게 관련되어 있음을 알 수가 있다.
따라서 본 연구에서 밝혀낸 춘계 베링해에서의 해양학적, 화학적 및 생물학적 요인들과 베링해에서 상업적으로 그리고 생태계적으로 중요 종인 명태의 분포에 관한 연구를 통하여 밝혀낸 결과를 토대로, 명태의 분포와 해양환경인자와의 관계를 구명하는데 중요한 자료로 활용할 수 있을 것이며, 또한 시험조사선에 의해 직접적인 조사를 통하여 밝혀낸 결과뿐만 아니라 인공위성 등 간접적인 정보기술을 이용하여 얻은 해양환경인자들로부터 명태의 분포를 예측할 수 있는 모델 개발에도 기초자료 로 이용되어질 수 있을 것이다. 앞으로 수산자원을 연구하는데 있어서 수산자원학적인 연구뿐만 아니라 해양환경학적인 연구도 병행하여 이루어져야 함을 강조하고 싶다.