Sixty-seven
장염비브리오(
장염비브리오는 수온이 17℃ 이상으로 상승하는 하절기에는 자유 유영의 형태로 해수에서 쉽게 검출되나 수온이 10℃ 이하로 낮아지면 해수에서의 검출빈도는 급격히 떨어지며 수온이 낮은 동절기에는 저질 또는 동물성 플랑크톤의 키틴질 등 에 부착하여 월동한다고 보고되어 있다(Kaneko and Colwell, 1975). 해수 및 어패류 등의 자연계에서 분리한 대부분의 장염 비브리오는 병원성 인자를 보유하고 있지 않은 비병원성 균주인 반면 식중독 사고의 환자 가검물에서 분리된 대부분의 장염 비브리오는 병원성 균주이다(Sakazaki et al., 1968; Honda and Iida, 1993). 이 균이 생산하는 대표적인 병원성 독소는 내열성용혈독(thermostable direct hemolysin, TDH), 내열성용혈독 관련용혈독(TDH-related hemolysin, TRH), serine protease 및 type III secretion systems (TTSS 1 및 2)를 통해 분비되는 각종 effector 단백질 등이 보고되어 있으나 정확한 병원성 메커니즘에 관한 설명은 아직도 많이 부족한 실정이다(Honda and Iida, 1993; Lee et al., 2002; Makino et al., 2003; Park et al., 2004; Kodama et al., 2007; Kodama et al., 2008).
페니실린 발견 이후 다양한 종류의 항생제는 사람과 동물의 치료, 질병예방 및 성장촉진 등의 목적으로 사용되고 있다. 2012년 농림수산검역검사본부(NVRQS, 2009) 에 의하면 돼지, 소 및 닭 등 축산용 항생제 판매량은 2005년 이래 서서히 감소하는 추세인데 비해 수산용 항생제 판매량은 2009년 178톤, 2010년 203톤 및 2011년 239톤으로 오히려 사용량이 소폭 증가하고 있는 것으로 파악되고 있다. 수산용 항생제 판매실적은 oxytetracycline, erythromycin, amoxicillin, 설파계열, ciprofloxacin, ampicillin 순으로 집계되고 있다(NVRQS, 2009). 지속적이고 다량의 항생제 사용은 양식어류에서 다양한 항생제에 내성을 나타내는 장염비브리오의 증가를 가중시키는 결과로 나타나고 있다(Son et al., 2005; Lee et al., 2007; Oh et al., 2008; Lee et al., 2009; Ryu et al., 2010; Yu et al., 2010).
세균이 항생제 내성을 갖게 되는 이유는 분해효소에 의한 항생제의 불활성화, 표적 항생물질의 변화, 세포막의 항생제 투과성 변화 및 세포 밖으로 항생제의 유출 등의 다양한 방법에 의한 것으로 알려져 있으며 이들 메커니즘이 단독 또는 복합적으로 작용하여 세균은 항생제에 내성을 갖게 된다. 획득내성은 세균 염색체의 유전자변이, 플라스미드(plasmid) 또는 트랜스포존(transposon)에 매개되는 내성유전자의 획득에 의해 생기며, 내성 유전자는 염색체 또는 plasmid DNA에 존재한다(Kuhl et al., 1978). 그람 음성 세균에서 항생제 다제내성 유전자를 encoding하고 있는 integron은 세균 염색체에서 이동성을 가진 DNA 단편인 transposon을 통하여 유전자의 한 복제단위에서 다른 복제단위로 이동되는데 가장 일반적인 방법은 접합을 통하여 동종 및 이종 세균으로 확산된다고 보고되어 있다(Rowe-Magnus and Mazel, 2002).
본 논문은 해수유래 장염비브리오의 각종 항균제 내성 양상 및 병원성 유전인자 보유성에 대한 기초자료를 수집하기 위하여 완도해역 해수에서 분리한 장염비브리오 67 균주를 대상으로 검토하였다. 또한 실험에 사용된 모든 균주에서 내성을 나타내는 ampicillin 및 oxacillin 항균제에 대해서는 최소발육억제 농도도 검토하였다.
실험에 사용한 장염비브리오는 2011년 6월부터 2012년 10월 까지 전남 완도군 군외면 영흥리 연안 표층 해수에서 분리한 67 균주 및 장염비브리오 RIMD2210633 (Makino et al., 2003)를 표준 균주로 사용하였다. 항균제 감수성 결과의 정도 관리를 위하여
실험에 사용한 DNA 증폭용 primers의 염기서열 및 증폭예상 DNA 크기 등은 Table 1에 제시하였다. Primers는 Bioneer (Daejon, Korea)에 의뢰 합성하였다. TDH, TRH,
[Table 1.] Primers used in this study

Primers used in this study
균주의 항균제 감수성은 Acar and Goldstein (1991)의 디스크 확산법과 미국 NCCLS (National Committee for Clinical Laboratory Standards, 2002)에 준하여 시험하였다. 식염 3% 첨가된 LB broth에 시험 균주를 접종하여 35℃에서 하룻밤 배양한 후 멸균생리식염수로 2회 세정하고 농도를 McFarland 0.5로 조정하여 Muller Hinton Agar (Merck, Germany) 평판에 도말 하였다. 여기에 검사 항균제 디스크를 고착하여 35℃에서 18시간 배양한 후 각 항균제에 의해 형성된 생육저지환의 크기를 측정하고 표준지표에 따라 감수성 여부를 평가하였다. 시험 항균제는 amikacin (AN; 30 μg), ampicillin (AM; 10 μg), cephalothin (CF; 30 μg), chloramphenicol (C; 30 μg), ciprofloxacin (CIP; 5 μg), erythromycin (E; 15 μg), gentamicin (GM; 10 μg), kanamycin (K; 30 μg), nalidixic acid (NA; 30 μg), oxacillin (OX; 1 μg), streptomycin (S; 10 μg), sulfamethoxazole/ trimethoprim (SXT; 23.75/1.25 μg), tetracycline (TE; 30 μg), trimethoprim (T; 5 μg), vancomycin (VA; 30 μg) 등 15종의 항균제 디스크를 사용하였다.
>
최소발육억제 농도(Minimum Inhibitory Concentration, MIC) 측정
시험 균주의 최소발육억제농도는 미국 NCCLS에 기초하여 변법으로 측정하였다. 멸균된 Muller Hinton broth (Merck, Germany)에 1 μg/mL에서 4,096 μg/mL까지 농도를 달리한 ampicillin 및 1 μg/mL에서 1,024 μg/mL까지 농도를 달리한 oxacillin을 첨가한 후 멸균된 소형시험관에 각 농도의 항생제가 첨가된 배지를 2 mL씩 분주하였다. 여기에 식염이 3% 첨가된 Luria-Bertani (LB) broth에서 하룻밤 전 배양한 시험균액 3 μL을 접종하여 35℃에서 18시간 정치 배양한 후 균 증식여부는 육안으로 확인하였다.
일반적으로 임상분리 장염비브리오의 대부분은 이 균의 대표적인 병원성 인자인 TDH 또는 TRH 유전자를 보유하고 있는 데 비해 해수 및 어패류 등의 환경유래 장염비브리오에는 이들 병원성 유전자 보유율은 매우 낮은 것으로 보고되어 있다 (Shirai et al., 1990). 그러나 새로운 검출방법의 개발로 인하여 환경 유래 장염비브리오에서도 병원성 유전자를 보유하고 있는 균주 검출율은 높아지고 있는 추세이다(Deepanjali et al., 2005; Jones et al., 2012; Ellingsen et al., 2013; Gutierrez et al., 2013). 해수 유래 장염비브리오의 항균성 내성 양상 및 병원성 유전자 보유성을 검토하기 위하여 2011년 6월부터 2012년 10 월까지 전남 완도군 군외면 영흥리 연안의 표층해수에서 API 20E Kit등을 이용하여 71균주의 장염비브리오를 분리하였다. 이들 균주를 대상으로 장염비브리오 동정을 위한 특이 유전자로 알려져 있는
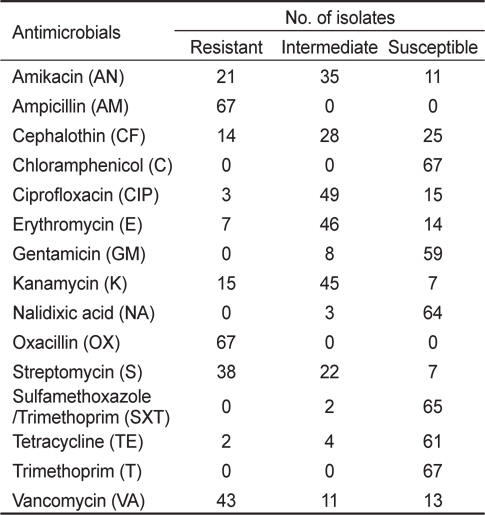
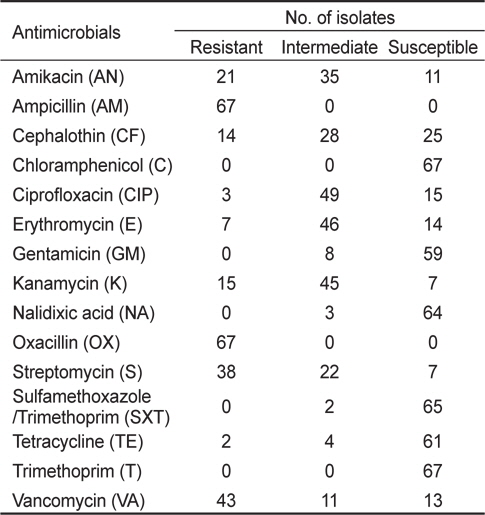
해수 및 어패류 등 다양한 환경시료에서 분리한 장염비브리오는 ampicillin, cephalothin, amikacin, cefoxitin, gentamicin, streptomycin, kanamycin, vancomycin 등의 단독 항균제에 내성을 나타낼 뿐만 아니라 다제 내성균의 검출빈도도 높은 것으로 보고되어 있다(Son et al., 2005; Lee et al., 2009; Ryu et al., 2010; Han et al., 2012). 완도해역 해수에서 분리한 장염비브리오 67균주를 대상으로 15종의 항균제에 대한 감수성 여부를 디스크 확산법으로 측정한 결과는 Table 2에 제시하였다. 15종의 항균제 중 10종의 항균제는 일부 또는 모든 균주에서 내성을 나타내었으며, chloramphenicol, gentamicin, nalidixic acid, sulfamethoxazole/trimethoprim 및 trimethoprim 등 5종의 항균제는 거의 모든 균주에서 감수성을 나타내었다. 특히, 실험에 사용된 67균주 전부는 ampicillin과 oxacillin에 내성을 나타내었다. 내성율이 높은 항생제 순서는 ampicillin과 oxacillin에 이어 43균주에서 내성을 나타내는 vancomycin (64.2%), 38균주에서 내성을 나타내는 streptomycin (56.7%), 21균주에서 내성을 나타내는 amikacin (31.3%), 15균주에서 내성을 나타내는 kanamycin (22.4%), cephalothin (20.9%), erythromycin (10.4%), ciprofloxacin (4.5%) 및 tetracycline (3.0%) 순서였다. 본 연구에 사용된 항균제 중 vancomycin은 그람 양성균에는 항균효능을 나타내나 그람 음성균에는 항균효능이 적기 때문에 내성을 나타내는 장염비브리오가 많은 것으로 사료된다.
Antimicrobial susceptibility and resistance of Vibrio parahaemolyticus isolated from surface seawater in Wando area
실험에 사용한 67균주는 chloramphenicol와 trimethoprim에는 100% 감수성을 나타내었으며, sulfamethoxazole/trimethoprim (97.0%), nalidixic acid (95.5%), tetracycline (91.0%), gentamicin (88.1%) 순으로 감수성을 나타내었다. 이 결과는 각종 항균제에 대한 장염비브리오의 내성 및 감수성 비율은 분리원, 분리시기 및 분리장소 등의 요인에 따라 차이가 있다는 기존의 연구결과와 거의 일치하는 경향이다(Son et al., 2003; Son et al., 2005: Lee et al., 2007; Lee and Park, 2010; Ryu et al., 2010; Han et al., 2012).
실험에 사용한 67균주에 대한 다제 항균제 내성에 관한 결과는 Table 3에 나타내었다. Ampicillin과 oxacillin의 2종의 항균제에 대해서 67균주에서 내성(100.0%)을 보이며, 여기에 1종의 항균제 즉 전체 3종의 항균제에 내성을 보이는 균주는 8주 (11.9%), 4종의 항균제에 내성을 보이는 균주는 26주(38.8%), 5종의 항균제에 내성을 보이는 균주는 19주(28.4%), 6종의 항균제에 내성을 보이는 균주는 4균주(6.0%) 및 7종의 항생제에 내성을 보이는 균주는 2균주(3.0%) 였다. 3종 또는 그 이상의 항균제에 대한 내성을 나타내는 다제 내성균은 전체 59균주 (88.1%)로 상당히 높은 비율을 차지하고 있었다(Table 3). 가장 높은 빈도의 항균제 내성조합은 AM-OX-VA-S로 14균주 (20.9%) 이며, 다음으로 AM-OX-VA-S-CF의 5균주(7.5%) 및 AM-OX-VA-S-K의 4균주(6.0%)로 파악되었으며 기타 항균제 내성조합 빈도는 3균주 이하로 매우 낮은 편이였다(Table 3).

Antimicrobial resistance patterns of Vibrio parahaemolyticus isolated from surface seawater in Wando area
>
장염비브리오의 ampicillin 및 oxacillin의 최소발육 억제농도 측정
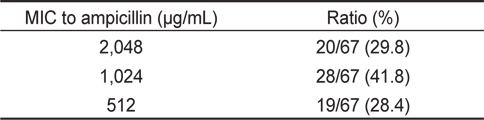
67균주에 대한 ampicillin 및 oxacillin에 대한 최소발육억제 농도(MIC)를 측정한 결과는 Table 4 및 5에 나타내었다. Table 4에 나타낸 바와 같이 ampicillin의 경우, 2,048 μg/mL의 MIC를 나타내는 균주는 20균주(29.8%), 1,024 μg/mL의 MIC를 나타내는 균주는 28균주로 전체의 41.8%를 차지하였다. 512 μg/ mL의 MIC를 나타내는 균주는 19균주(28.4%)였다. 실험에 사용한 장염비브리오 67균주의 ampicillin에 대한 최소발육억제 농도의 평균치는 1,184 μg/mL로 확인되었다(Table 4). 이 결과는 자연계에서 분리한 장염비브리오는 일반적으로 ampicillin에 고도내성을 나타내고 있다는 기존의 결과와 대체로 유사하다(Tanil et al., 2005; Lee et al., 2007; Ferrini et al., 2008; Lee et al., 2009; Lee and Park, 2010; Lee et al., 2011).
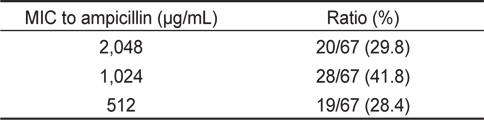
Minimum inhibitory concentration (MIC) to ampicillin of Vibrio parahaemolyticus isolated from surface seawater in Wando
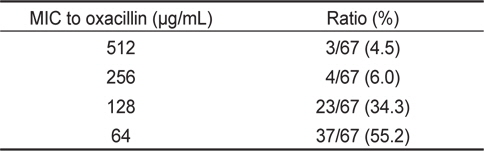
또한 oxacilllin 에 대한 최소발육억제농도를 측정한 결과, 512 μg/mL의 MIC를 나타내는 균주는 3균주(4.5%), 256 μg/mL의 MIC를 나타내는 균주는 4균주(6.0%), 128 μg/mL의 MIC를 나타내는 균주는 23균주(34.3%) 및 64 μg/mL의 MIC를 나타내는 균주는 37균주(55.2%)로 최소발육억제농도의 평균치는 117.5 μg/mL로 확인되었다(Table 5). 장염비브리오의 oxacillin에 대한 최소발육억제농도는 ampicillin에 비해 약 10배 정도 낮은 농도인 것으로 확인되었다.
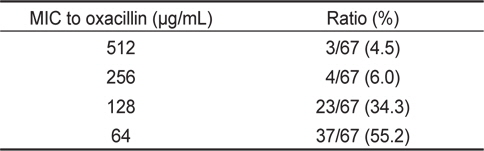
Minimum inhibitory concentrations (MIC) to oxacillin of Vibrio parahaemolyticus isolated from surface seawater in Wando area
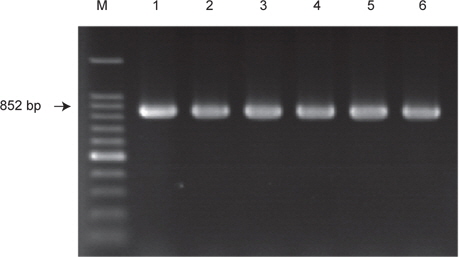
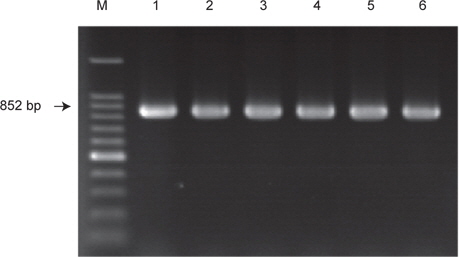
장염비브리오의 ampicillin 내성유전자 VPA0477 (β-lactamase)의 존재유무를 PCR assay로 확인한 결과 실험에 사용한 모든 균주에서 양성으로 확인되었으며, 일부 균주에 대한 PCR assay 결과는 Fig. 2에 나타내었다. 이 결과는 장염비브리오에 존재하는 VPA0477 유전자는 장염비브리오 동정을 위한 표적유전자로 활용될 가능성이 제기된다.
본 실험에 제공된 완도해역에서 분리한 67균주 장염비브리오는 모든 균주에서 ampicillin 및 oxacillin에 내성을 나타낼 뿐만 아니라 다수의 항균제에 대해서도 내성 보유현상이 보편화되어 있다는 점에서 장염비브리오의 항균제 내성에 관한 꾸준한 모니터링 및 감시가 필요하다고 판단된다. 또한 다양한 항균제 내성 유전자의 동정 및 염색체 DNA상에서의 존재양상 등의 파악은 내성유전자의 획득 및 확산 메커니즘을 이해하는데 큰 도움이 될 것으로 기대된다.