The burrowing cricket genus
In East Asia, around the Korean peninsula, the numbers of
In the course of taxonomic studies on Korean crickets and its sound recording projects, the following results were produced relating to the genus
On the other hand,
In the case of
The aim of this paper is to review the burrowing cricket genus
Order Orthoptera Olivier, 1789
Family Gryllidae Laicharting, 1781
Subfamily Gryllinae Laicharting, 1781
Genus Velarifictorus Randell, 1964
Type species:
Diagnosis. Body coloration dark brown, appendages more brightly mottled. Sexual dimorphism remarkable: male with enlarged round head and full winged tegmina, but female with relatively normal head and short tegmina. Head with transverse strip between lateral ocelli, but sometimes not noticeable. Hind wings degenerated, not surpassing tegmina. Fore tibia with only outer tympanum. Male epiphallus with a pair of prominent lateral lobes and a wide median lobe.
Remarks. It should be noted that Randell (1964) designated
Subgenus Velarifictorus Randell, 1964
Type species:
Diagnosis. Male epiphallus with relatively long, but not high hind median projection directed backwards. The other two subgenera diagnoses are as follows:
Remarks. All known Korean members belong to the nominate subgenus.
1*Velarifictorus aspersus borealis Gorochov, 1985)
(Tables 1, 2, Figs. 1A, B, 2A, 3A, 4A, 5A
Velarifictorus aspersus borealis Gorochov, 1985: 91 (type locality: China, Beijing); Storozhenko and Paik 2007: 100; Paik et al., 2010: 78.
Gryllus aspersus Walker, 1869: 39 (type locality: Hongkong); Scapsipedus aspersus: Wu, 1935: 70; Cho, 1959: 144; 1969: 769; Choo and Choi, 1983: 49; Song, 1988: 357; 1989: 283; Bae and Lee, 1990a: 59; 1990b: 67; Velarifictorus aspersus: Kwon and Huh, 1994: 49; Moon and Yoon, 1994: 106; Kwon et al., 1996: 104; Hua 2000: 56; Paek et al., 2010: 35; Kim, 2011: 65.
=Gryllodes berthellus Saussure, 1877: 205 (type locality: Japan, Amboine, Banka, China); Shiraki, 1930: 218 (“Keijo” =Seoul); Doi, 1932: 38; Ju, 1993: 286; Gryllodinus berthelles [!]: Ju, 1969: 19; Gryllodes berhellus [!]: Hua, 2000: 54 [a synonym by Chopard, 1967].
=Scapsipedus mandibularis Saussure, 1877: 246 (type locality: India, Bombay, Japan); Doi, 1932: 38; Cho, 1959: 145; 1969: 769; Bae and Lee, 1990a: 59; 1990b: 67; Kwon and Huh, 1994: 49; Moon and Yoon, 1994: 106; Kwon et al., 1996: 104; Scapsipedlus [!] mandibularis: Ju, 1969: 21; Velarifictorus mandibularis: Hua, 2000: 57 [a synonym by Chopard, 1967].
Material examined (50 specimens). Korea: Gyeongsangnam- do: 1♂ (light trap), Jinju-si, Yugok-dong, Nabul-ri, 25-26 Sep 1998, Byeon JH; 14♂8♀, ibid. 16 Aug 2012, Kim TW, Kim OS; 1♂1♀, Sacheon-si, Chukdong-myeon, Baechun-ri, 17 Aug 2012, Kim TW; 1♂, Tongyeong-si, Jeongryang-dong, 8 Oct 2011, Sung KS; 2♂, Tongyeong-si, Inpyeong-dong, Gyeongsang National University College of Marine Science Campus, 23 Aug 2006, Jang YK; 5♂8♀, ibid. 17 Aug 2012, Kim TW, Kim OS; Jeollanam-do: 5♂ 3♀, Yeosu-si, Sangam-dong Sangam elementary school, Mt. Yeongchuisan, 28 Aug 2012, Kim OS; Japan: 1♂, Kyoto Prefecture, Kyoto City, Sakyo-ku, Yoshida, Mt. Yoshidayama, alt. 100-120 m, 21 Sep 1988, S. Fujii.
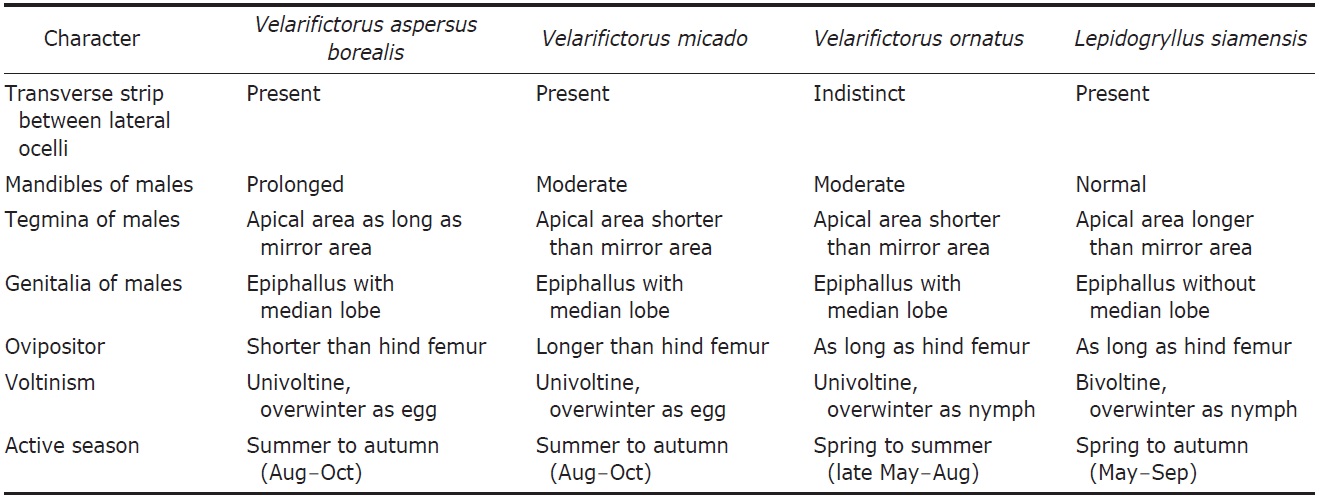
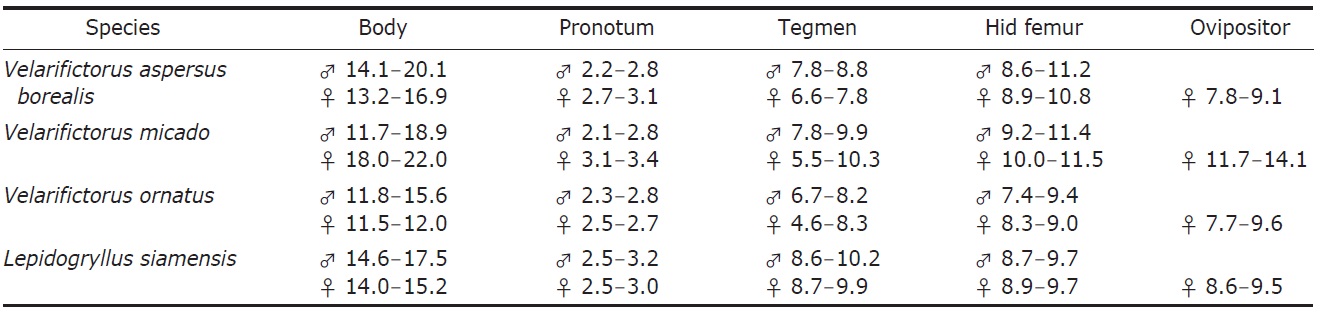
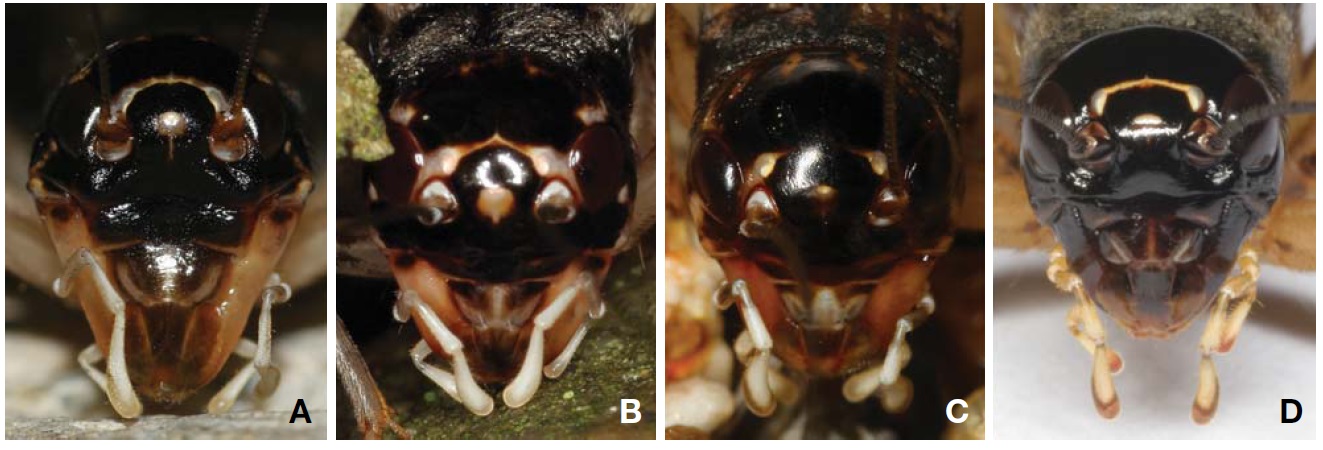
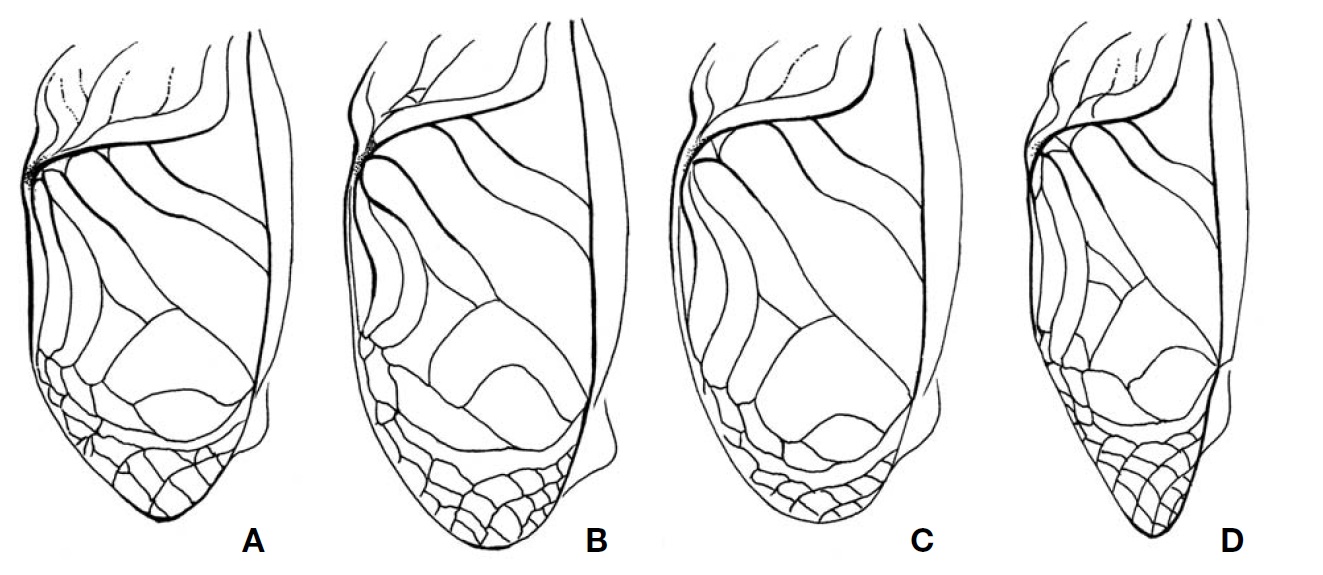
Description. Male (Fig. 1A): Head globulous, sufficiently wider than anterior margin of pronotum in dorsal view; vertex transversely dark brown; occiput bright brown with six longitudinal pale strips posteriorly; a distinct strip transverse between lateral ocelli, its length two times wider than horizontal diameter of antennal socket; median ocellus located between inner upper margins of antennal sockets, surrounded with a short bright line linked downwardly; subocular area weakly depressed, coarsely furrowed; mandibles elongated, thurs clypeal suture transverse higher than mid in head, the suture thickened in upper margins, but diminished in mid; scape as long as wide; palpi bright. Pronotum a little shorter than fore femur, two times wider than long in dorsal view; anterior margin slightly wider than posterior one, both truncated, pilose; lateral lobe of pronotum slightly deeper than pronotal length, lower half area bright. Tegmina (Fig. 3A) rather wider than posterior margin of pronotum, not reaching abdominal end; harp veins two; chordal veins three; mirror area divided, 1.2 times wider than long; apical area well netted, as long as mirror area; lateral field deeper than lateral lobe of pronotum, with 5-6 longitudinal veins. Fore tibia with largely oval-shaped outer tympanum, 3.5 times longer than wide; inner tympanum vestigial, not discernable. Hind femur 2.7 times longer than middle femur. Hind tibia with five pairs of large dorsal spines on both sides, the largest inner apical spur as long as half of basitarsus. Hind barsitarsus two times longer than remaining two tarsomeres combined length; six dorsal spinules on both sides; inner apical spur as long as last tarsomere. Cercus as long as three fifths of hind femur. Subgenital plate as long as half of basal width, its hind margin weakly notched in mid.
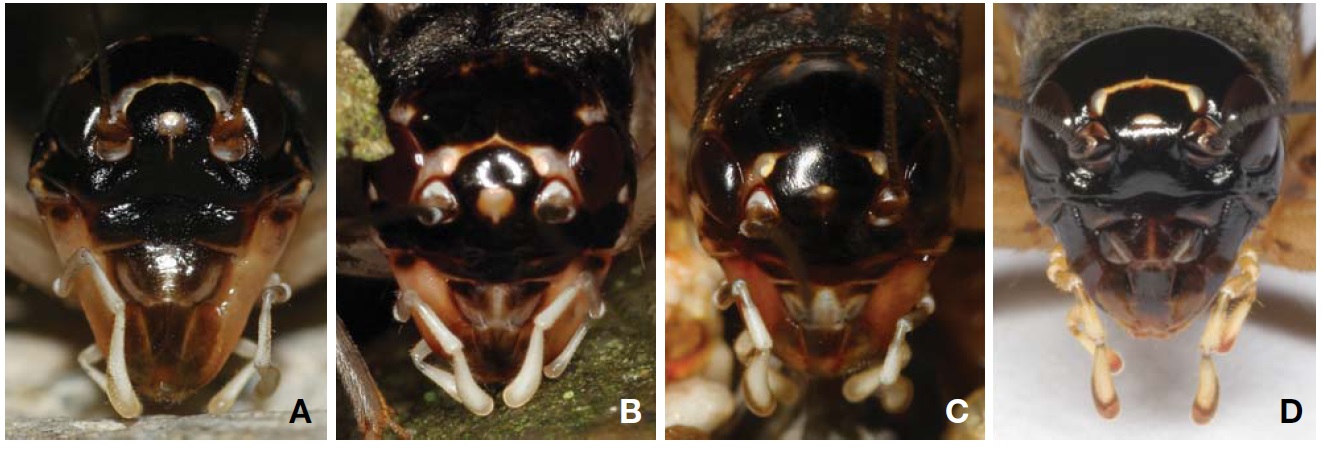
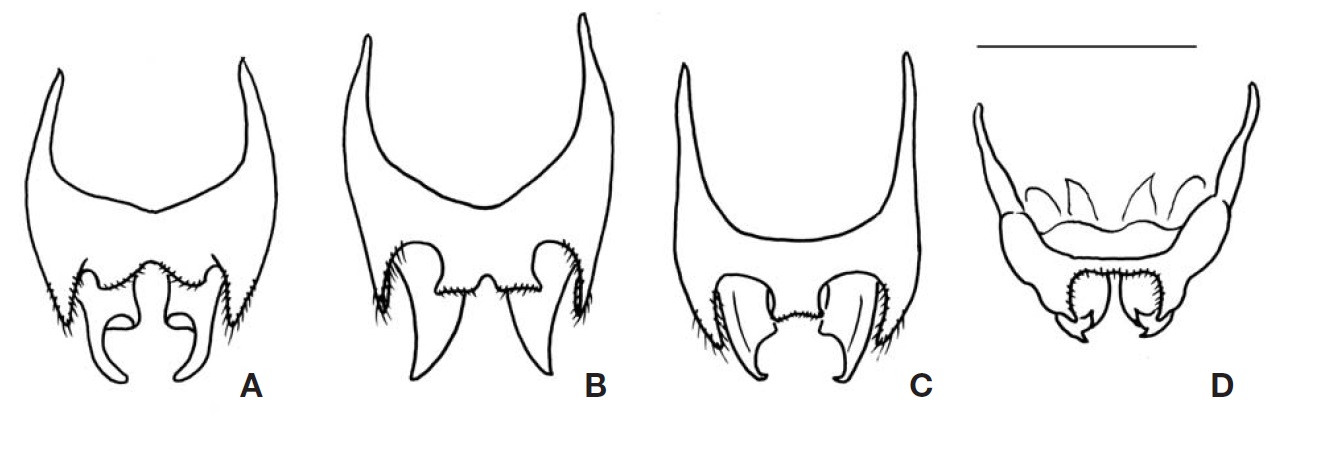
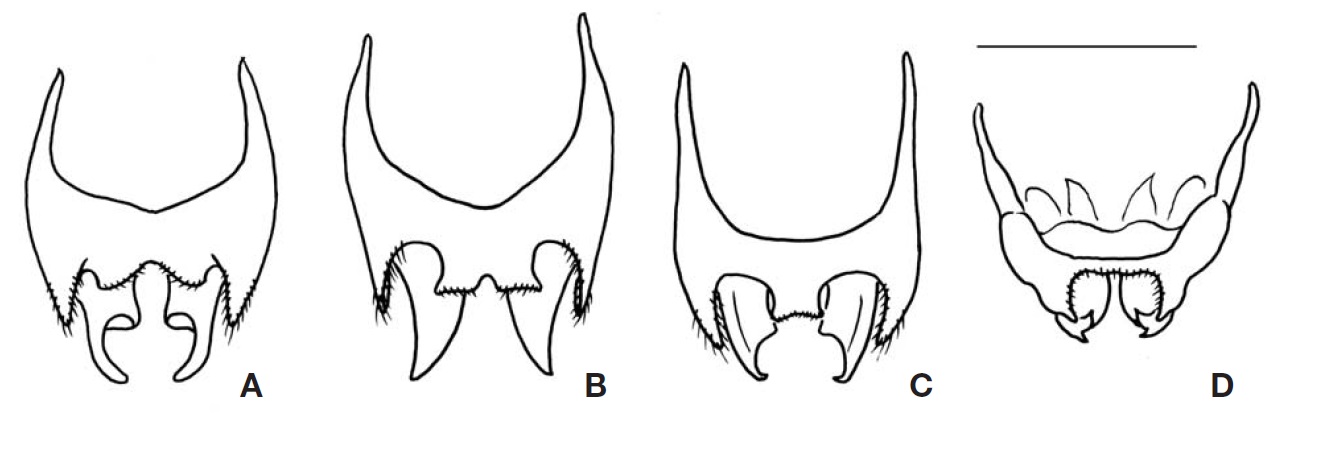
Genitalia (Fig. 4A): Epiphallus with a broad and short median lobe, but which is rather distinctly divided posteriorly as a pair of small obtuse-angular lobes by median concavity; lateral lobes longer than middle lobe, with long hairs. Ectoparamere greatly projected beyond hind margin of epiphallus, its basal half rather thickened, but its distal half slightly incurved and sharply tapered at apex, hook-shaped.
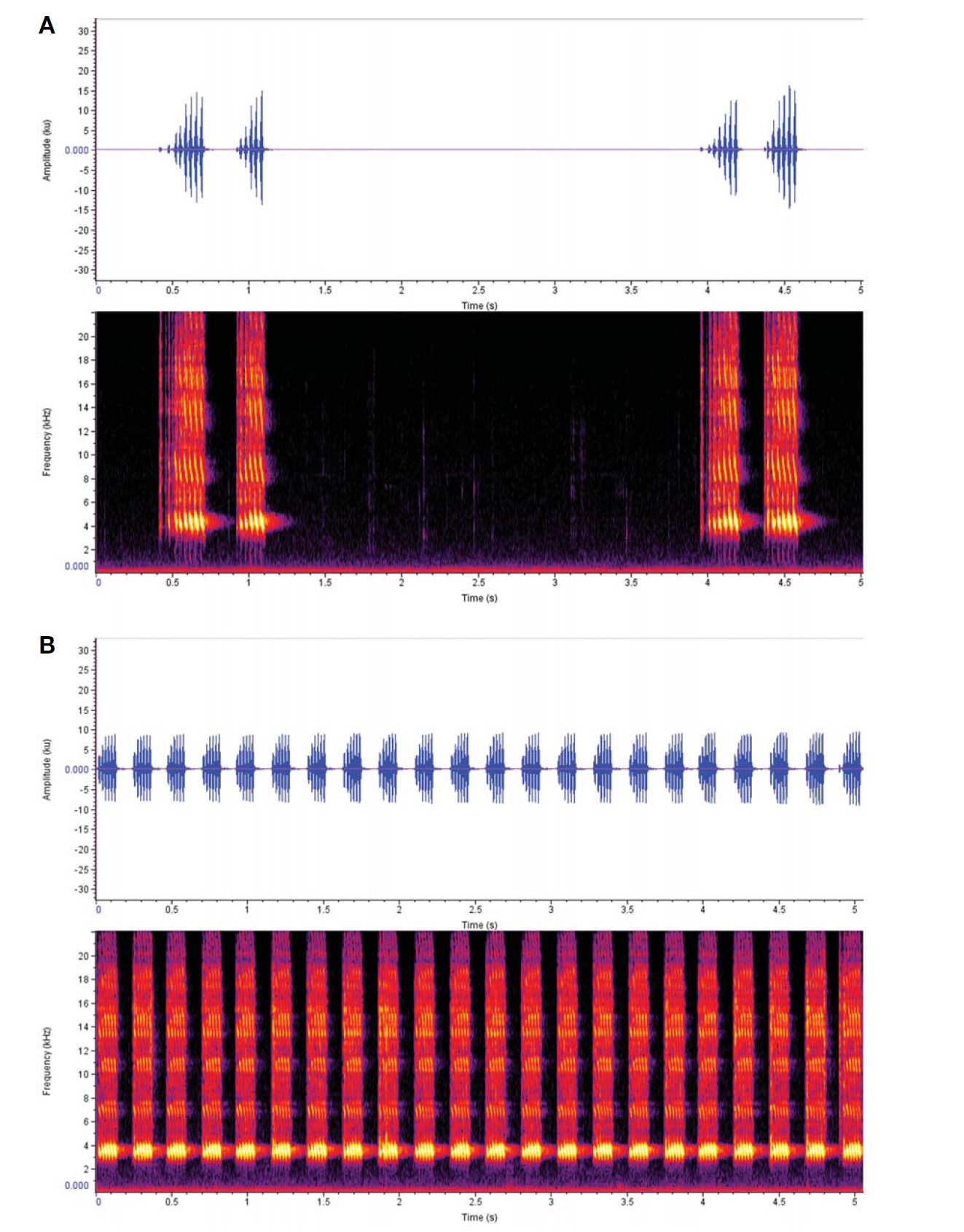
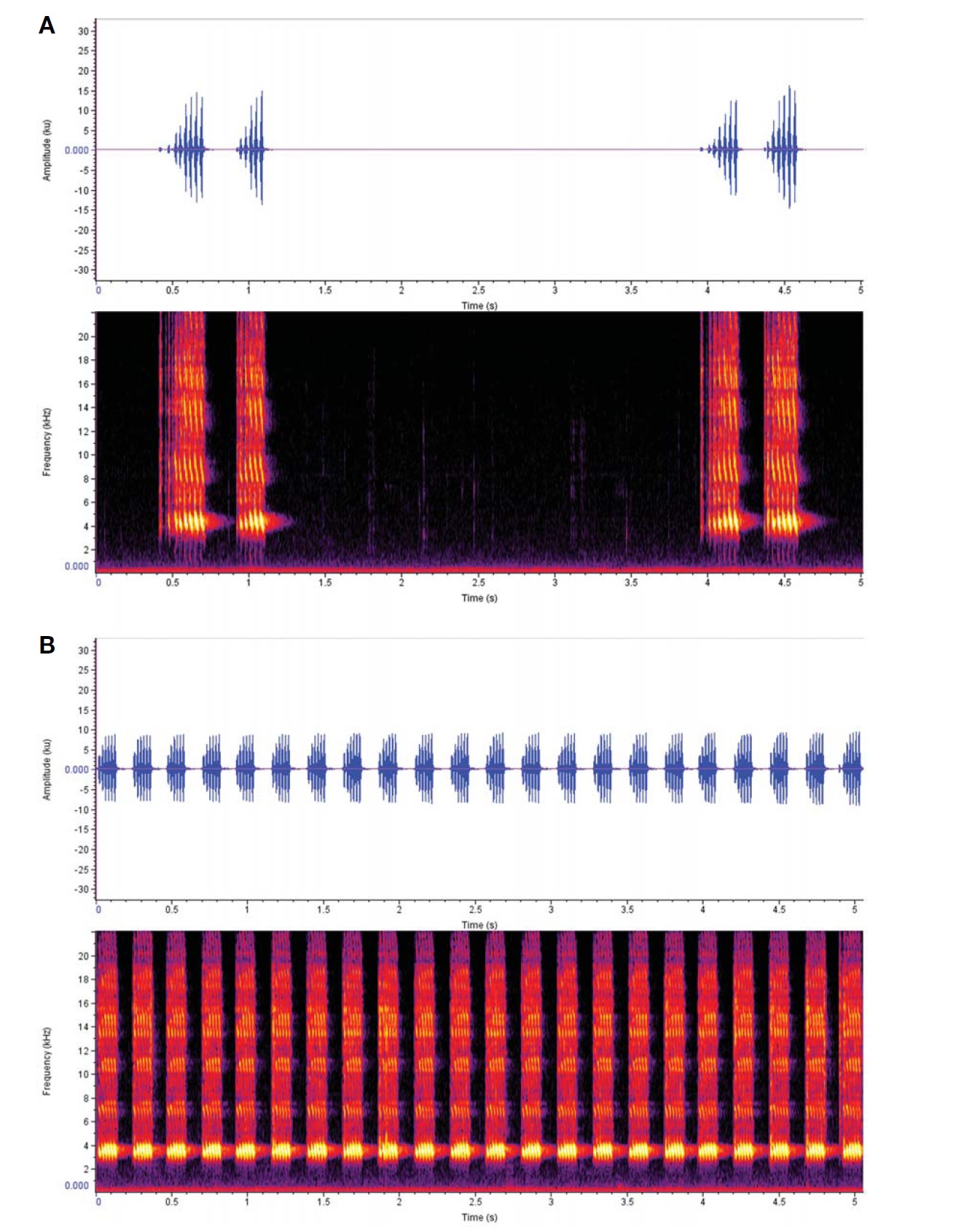
Calling sound (Fig. 5A): Shortly and slowly repeated song phrase, each consisting of 1-3 distinctive chirps; their intervals sometimes irregular depending on outer conditions, when regularly repeated mean chirps two; a chirp with undulating 5-8 syllables at 29° condition; each syllable lasted 0.03 s; dominant frequency spectrum 3.5-5 KHz.
Female (Fig. 1B): Head smaller than in male, mandibles moderate, not enlarged. Tegmina shorter than male, just covering two thirds of dorsal abdomen, its apex roundly truncated; medial area longitudinally bright. Subgenital plate convex, roundly trapezoidal, its hind margin narrowly truncated. Ovipositor as long as hind femora.
Distribution.
Remarks. Although there were plentiful records of
1*Velarifictorus (Velarifictorus) micado (Saussure, 1877) (Tables 1, 2, Figs. 1C, D, 2B, 3B, 4B, 5B)
Scapsipedus micado Saussure, 1877: 247 (type locality: Japan, Celebes); Velarifictorus micado: Kwon and Huh, 1994: 50; Kwon et al., 1996: 104; Paik et al., 2010: 30, figs. 36ab; Kim, 2011: 67; Velarifictorus mikado [!]: Storozhenko and Paik, 2007: 100; Paek et al., 2010: 35.
Material examined (147 specimens). Korea: Gangwon-do: 1♂3♀ (light trap), Gangneung-si, Samsan-ri, Buyeongyegok Valley, 18-19 Aug 2001, Kim JI, Kim SY; 1♂5♀, Gangneung- si, Okgye-myeon, Sangye-ri, Mt. Seokbyeongsan, 27-28 Aug 2002, Kim TW; 1♂3♀, Yeongwol-gun, Gonggiri, Mt. Sambangsan, 17-18 Sep 2001, Kim TW, Kim MA; 1♀ (light trap), Yeongwol-gun, Mt. Baekdeoksan, Gwaneumsa Temple, 26 Aug 2001, Kim SY, Kim AY; 1♀, Jeongseongun, Nam-myeon, Jeungsan, 19 Sep 2004, Kim TW; 3♀, Chuncheon-si, Gangchon, 20 Sep 1996, Huh R; 1♂, Hwacheon- gun, Haesanryeong Pass, 3 Oct 2006, Kim TW; 1♂, Cheongsong-gun, Chongpyeong, 16 Aug 1988, Oh HJ; 1♀, Wonju-si, Haenggu-dong, Mt. Chiaksan, Hwanggol Office, 1 Sep 2006, Kang TH; Gyeonggi-do: 5♂6♀ (pitfall), Gapyeong- gun, Mt. Cheonggyesan, 27 Sep 2002, Jeong MP; 1♂, Incheon-si, Isl. Boleumdo, 25 Aug 2007, Kim TW; 1♂, Goyang-gun, Madu-dong, 14 Aug 1994, Park SK; 1♂, Goyang- gun, Hwajeong-dong, 1 Sep 2009, Kim TW; 4♂1♀, Yeoncheon-gun, Jeonguk-eup, Hantangang River-bed, 23 Sep 2011, Kim TW; 1♂, Pocheon-gun, Gwangneung Arboretum, 12 Sep 1991, Jeong SM; 1♀, ibid. 6 Aug 1992, Park SJ; 1♀, Gwangju-si, Eommi-ri, 15 Sep 1984, Kim JI; 1♂, Gunpo-si, Mt. Surisan, 7 Oct 2001, Kim TW; 1♂1♀, Pajusi, Mt. Gamaksan, 8 Sep 1984, Kim JI; 1♀, Namyangju-gun, Mt. Cheonmasan, 21 Sep 1987, Kim MS; 1♀, Suwon-si, Cheongcheon-dong, 9 Oct 1987, Lee JH; 2♀, Ansan-si, Daebudo, Sihwaho Lake, 27 Aug 2004, Kim TW; 2♂3♀ (pitfall), Ansan-si, Singil-cheon, 19 Oct 2002, Jeong MP; 6♂7♀ (pitfall), Hanam-si, Misa-dong, 28 Sep 2002, Jeong MP; 1♀, Uijeongbu-si, Gosan-dong, 23 Aug 1992, Choi EA; 1♀, Mt. Bulamsan, 18 Sep 1982, Lee MK; 1♀, Yangpyeong-gun, Yeonsu-ri, 31 Aug 1980, Chang KS; Seoul: 2♂1♀, Eunpyeong- gu, Mt. Bukhansan, 1 Oct 2001, Kim TW; 2♂1♀, ibid. 3 Oct 2003, Kim TW; 2♀, Bukhansanseong, 26 Sep 1991, Jeong SM; 1♀, Jungryang-gu, Junghwa-dong, 12 Sep 1988, Sim HJ; 1♀, Jungryang-gu, Mt. Mangwusan, 4 Oct 1996, Lee SY; 1♀, Uenpyeong-gu, Yeokchon-dong, 9 Oct 1987, Lee MK; 1♀, Dongjak-gu, Sangdo-dong, 4 Aug 1985, Han SO; 1♀, Gangnam-gu, Mt. Cheonggyesan, 14 Aug 1985, Yoon HJ; 1♂, Mt. Kachisan, 25 Aug 1982, Lee EK; 2♂, Mt. Gwanaksan, 25 Sep 1982, Yoo YS, Yoo HK; 1♂, ibid. 8 Sep 1990, Seo MJ; 1♂, Seocho-gu, Mt. Woomyeonsan, 1 Sep 1991, Oh JM; 1♀, Wolgye-dong, 6 Nov 1982, Park HM; 1♀, Seongbuk-gu, Dongseon-dong, 30 Sep 1997, Cho KH; 1♂, Mok 3-dong, 22 Aug 1996, Jeong EJ; 1♂, Dobong-gu, Sinchang-dong, 24 Aug 1987, Park K; Chungcheongbuk- do: 1♀ (light trap), Jecheon-si, Deoksan-myeon, Dogi-ri, Mt. Wolaksan, Munsubong, 14-15 Sep 1996, Jeon JS; 3♀, Chungju-si, Sancheok-myeon, Mt. Cheondeungsan, 28 Aug 2002, Kim TW; 2♀, Chungju-si, Jongmin-dong, Mt. Gyemyeongsan, 29 Aug 2002, Kim TW; 4♂1♀, Chungjusi, Gageum-myeon, Chungju Expressway Resting Place (a down line), 25 Aug 2011, Kim TW; Chungcheongnam-do: 1♀, Gongju-si, Jakgol, 8 Aug 1982, Kim HK; 1♀, Geumsangun, Chubu-myeon, Sundang-ri, Mt. Seodaesan, 28 Sep 2002, Kim AY; 1♂, Hongseong-gun, Gwangcheon-eup, 19 Sep 1998, Yoo JS; Gyeongsangbuk-do: 1♀, Daegu-si, Nam-gu, Daemyeong-dong, 18 Aug 1995, Park EM; Gyeongsangnamdo: 1♀ (light trap), Goseong-gun, Sangri-myeon, Mt. Muisan, Munsuam, 14-15 Oct 1999, An TH; 1♂ (light trap), Namhae- gun, Simcheon-ri, Simcheoncheon, 29-30 Aug 1998, Jeon JS; 2♂1♀, Sacheon-si, Chukdong-myeon, Baechunri, 13 Sep 2001, Paik YH; 1♂ (light trap), Sacheon-si, Waryong- dong, Mt. Waryongsan, Cheongryongsa, 5-6 Oct 1996, An TH; 1♀ (light trap), Jinju-si, Naedong-myeon, Sinyul-ri, 22-23 Sep 1998, Seol JH; 1♂ (light trap), Jinju-si, Ihyeondong, Mt. Sukhosan, 23-24 Sep 1998, Jeon JS; 1♂2♀, Jinju-si, Yugok-dong, Nabul-ri, 16 Aug 2012, Kim TW, Kim OS; 1♂, Tongyeong-si, Jeongryang-dong, 8 Oct 2011, Sung KS; 1♀, Hadong-gun, Hwagae-myeon, Ssanggyesa Temple, 1 Sep 2001, Kim TW; 1♂, ibid. 14 Sep 2010, Kim TW; 1♀, Busan-si, Yeongdo-gu, Cheongsan APT, 30 Aug 2000, Kim SK; 1♂4♀, Haman-gun, Mt. Yuhangsan, 21-23 Sep 2006, Lee DH, Park JH, Park YN; 1♀, Busan-si, Gijang-gun, Ilgwang- myeon, Won-ri, Mt. Dareum, 20 Sep 2008, Sang HS; 1♀, Busan-si, Gijang-gun, Gijang-eup, Mt. Samgak, 19 Sep 2008, Moon TY; Jeollabuk-do: 3♂1♀ (pitfall), Buan-gun, Haseo-myeon, Haechang-ri, 24 Sep 2001, Jeong MP; Jeollanam- do: 1♀, Mokpo-si, Sanjeong-dong, Samhakdo, 16 Oct 2004, Kim TW; 1♂4♀, Gwangyang-si, Dapgok, Mt. Baekunsan, 15 Sep 2001, Paik JC; 1♂1♀, Suncheon-si, Maegok- dong, 3 Oct 2007, Paik JC; Jeju-do: 1♂, Namjeju-gun, Andeok-myeon, Sagye-ri, Mt. Sanbangsan, 14 Aug 2004, Kim JK, Kim TW; 1♂, Namjeju-gun, Andeok-myeon, Andeokgyegok Valley, 15 Oct 2008, Kim TW; 1♂, Jeju-si, Nohyeong-dong, 7 Oct 2006, Jung SH; 1♂, Seoguipo-si, Cheonjiyeon-pokpo, 10 Oct 2011, Kim TW.
Description. Male (Fig. 1C): Head globular, slightly wider than anterior margin of pronotum; vertex roundly merging with frons; occiput with six pale longitudinal strips posteriorly; a distinct strip transverse between lateral ocelli, merging with ∧-shaped ecdysial line, its length two times wider than horizontal diameter of antennal socket; median ocellus located between inner upper margins of antennal sockets, surrounded by a bright line linked downwardly; subocular area weakly depressed, coarsely punctured; mandibles vertical; clypeal suture transverse lower than mid in head; scape as long as wide; palpi bright. Pronotum as long as fore femur, two times wider than long in dorsal view; anterior and posterior margins truncated, pilose; lateral lobe of pronotum as deep as pronotal length, lower margin bright. Tegmina (Fig. 3B) slightly wider than hind margin of pronotum, not reaching abdominal end; harp veins two; chordal veins tree; mirror area divided, as long as wide; apical area well netted, slightly shorter than mirror area; lateral field deeper than lateral lobe of pronotum, with 6-7 longitudinal veins. Fore tibia with largely oval-shaped outer tympanum, its length four times longer than wide; inner tympanum vestigial, not discernable. Hind femur three times longer than middle femur. Hind tibia with five pairs of large dorsal spines on both sides, the largest inner apical spur as long as a half of basitarsus. Hind barsitarsus two times longer than remaining two tarsomeres combined length; 5-6 dorsal spinules on both sides; inner apical spur as long as two thirds of last tarsomere. Cercus as long as three fifths of hind femur. Subgenital plate as long as half of basal width, its hind margin weakly notched in mid.
Genitalia (Fig. 4B): Epiphallus with a moderate-sized median lobe, its hind margin rather transversely truncated, a small emargination present on mid posteriorly, but sometimes not discernable; lateral lobes longer than middle lobe, with long hairs. Ectoparamere acute blade-shaped, greatly projected surpassing hind margin of epiphallus, its apex triangularly pointed.
Calling sound (Fig. 5B): Continuously lasting song phrase, distinctive chirps 4-5 times regularly repeated for 1 s; a chirp with undulating 6-8 syllables at 27° condition; each syllable lasted 0.02 s; dominant frequency spectrum 3-4 KHz.
Female (Fig. 1D): Head slightly smaller than in male, without subocular depression. Tegmina shorter than male, just covering two thirds of abdomen, its apex narrowly rounded; medial area longitudinally bright. Subgenital plate convex, roundly triangular, its hind margin narrowly truncated. Ovipositor 1.5 times longer than hind femora.
Distribution. Korea, Japan, China, Vietnam, Sulawesi; North America (introduced).
Remarks. This burrowing cricket is the most widespread
1*Velarifictorus (Velarifictorus) ornatus (Shiraki, 1911)
(Tables 1, 2, Figs. 1E, F, 2C, 3C, 4C, 5C)
Material examined (38 specimens). Korea: Gyeonggi-do: 1♀, Gwangju-si, Gwangju-eup, 13 Jun 1992, Shin MJ; 1♂, Gunpo-si, Mt. Surisan, 19 Jun 2005, Kim TW; 1♀, Yonginsi, Giheung, 31 May 2003, Choi JE; 1♀, Incheon-si, Ongjingun, Deokjeok-myeon, Seopo-ri, 14 Jun 1962, Na YJ; 1♀, Incheon-si, Juan-dong, 14 Jun 1969, Kim MR; 1♂, Namyangju- si, Mt. Cheonmasan, 14 Jun 1979, Kim TS; 2♀, Hwaseong- si, Jangan-myeon, Bangchukmal, 16 Jul 2009, Park YK; 2♀, Paju-si, Beopwon-eup, 6 Jul 2008, Kim TW; 1♂, Paju-si, Gwangtan-myeon, Bogwangsa Temple, 9 Jun 1979, Kim SH; Seoul: 1♀, Daebang-dong, 31 May 1963, Kim HJ; 1♀, Seongbuk-gu, Jeongneung, 21 Jun 1959, Kim YI; 1♂, ibid. 13 Jun 1959, Lee SA; 1♀, Sinchon, 26 May 1965, Shin SJ; 1♂, Eunpyeng-gu, Bulgwang-dong, 28 Jun 1960, Hong HJ; Chungcheongnam-do: 2♂3♀, Seosan-si, Haemi-myeon, Hanseo University Campus, Mt. Gayasan, 22 Jun 2001, Kim TW; Gyeongsangbuk-do: 1♂, Sangju-si, 23 Jun 2000, Park JK; Gyeongsangnam-do: 1♀ (light trap), Goseong-gun, Sangri- myeon, Yongtae-ri, Mt. Sanseongsan, 10-11 Aug 1996, Jeon JS; 1♂1♀, Hadong-gun, Hwagae-myeon, Mt. Jirisan, Ssanggyesa Temple, 31 Jun 2001, Kim TW; 2♀, Geoje-si, Yeoncho-myeon, Songjeong-ri, 11 Jun 2009, Kim TW; 1♀ (sweeping), Yangsan-si, Mt. Wonhyosan, 28 Aug 2002, Do YH; Jeollanam-do: 3♂, Wando-gun, Wando-eup, Wando Tower, 1 Jul 2010, Kim TW; 1♂, ibid. 19 Jul 2012, Kim TW; 2♂1♀, Wando-gun, Cheongsan-myeon, Yeoseo-ri, 20 Jul 2012, Kim TW; 1♀, Gokseong-gun, 7 Jul 1999, Paik YH; 1♂, Haenam-gun, Munnae-myeon, Jindo Bridge, 16 Jun 2011, Sung KS; 1♂, Jindo-gun, Dongoe-ri, Mt. Suribong, N34°28′37.8′′, E126° 18′04.1′′, alt. 183 m, 29 Jun 2004, Sohn JC, Park HJ, Nam SC, Han YE.
[Table 1.] Differential characters of Velarifictorus and Lepidogryllus in Korea
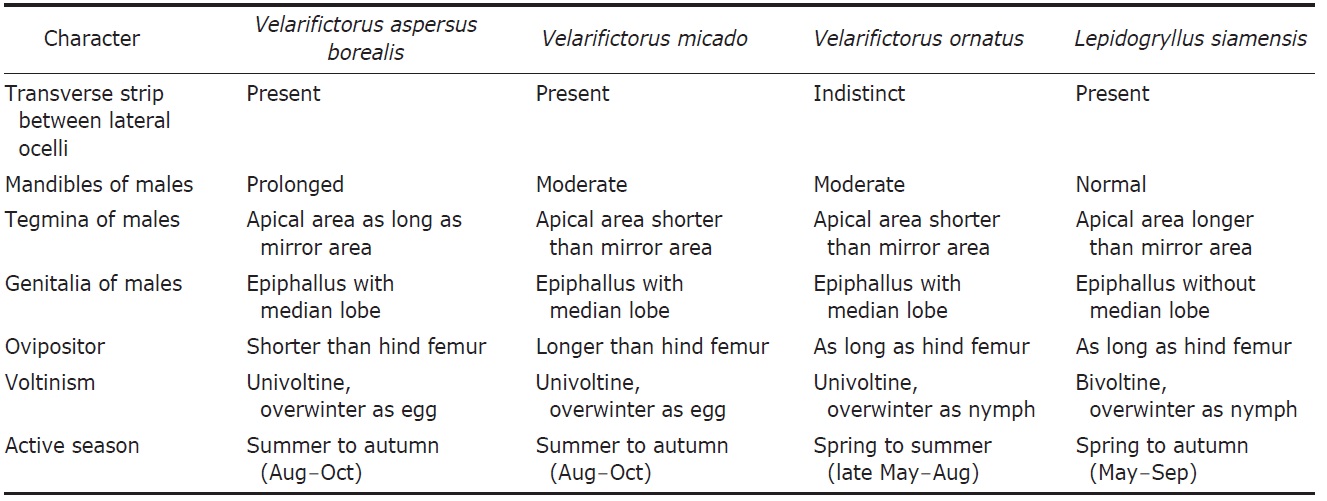
Differential characters of Velarifictorus and Lepidogryllus in Korea
[Table 2.] Measurements of Velarifictorus and Lepidogryllus in Korea (length in mm)
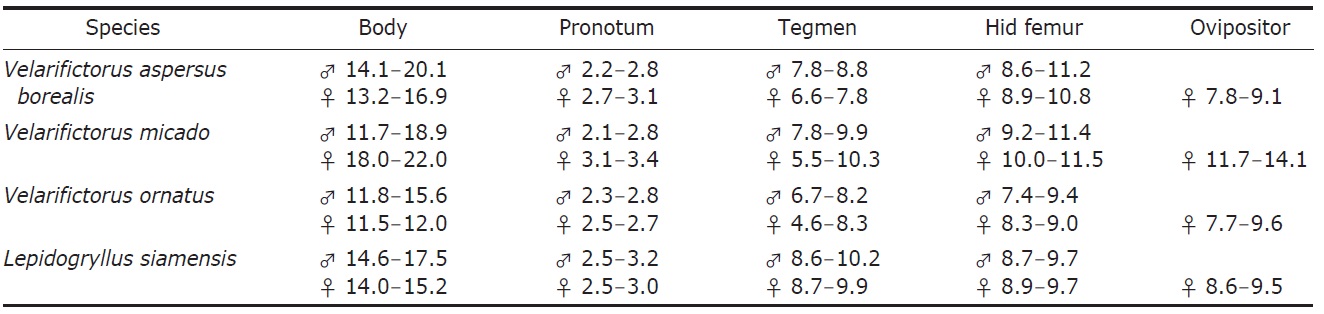
Measurements of Velarifictorus and Lepidogryllus in Korea (length in mm)
Description. Male (Fig. 1E): Head globular, slightly wider than anterior margin of pronotum; vertex and frons roundly merging; occiput with six pale longitudinal strips posteriorly; a transverse strip absent between lateral ocelli, diminished to feeble ecdysial line, thurs hardly distinguished; median ocellus located between inner upper margins of antennal socket, linked with short bright line downwardly; subocular area indistinctly furrowed; mandibles vertical; clypeal suture transverse lower than mid in head; scape as long as wide; palpi bright. Pronotum as long as fore femur, two times wider than long in dorsal view, with a pair of dorsal crescent patterns; anterior and posterior margins truncated, pilose; lateral lobe of pronotum as deep as pronotal length, lower half area bright. Tegmina (Fig. 3C) as wide as hind margin of pronotum, not reaching abdominal end; harp veins two; chordal veins three; mirror area divided, as long as wide; apical area well netted, as long as two thirds of mirror area; lateral field deeper than lateral lobe of pronotum, with 5-6 longitudinal veins. Fore tibia with largely oval-shaped outer tympanum, its length three times longer than wide; inner tympanum vestigial, not discernable. Hind femur 2.7 times longer than middle femur. Hind tibia with five pairs of large dorsal spines on both sides, the largest inner apical spur as long as a half of basitarsus. Hind barsitarsus 1.7 times longer than remaining two tarsomeres combined length; 5-6 dorsal spinules on both sides, inner apical spur as long as two thirds of last tarsomere. Cercus as long as thee fifths of hind femur. Subgenital plate as long as half of basal width, its hind margin weakly notched in mid.
Genitalia (Fig. 4C): Epiphallus with relatively small and short median lobe, its hind margin transversely truncated, trapezoidal; lateral lobes longer than middle lobe, with long hairs. Ectoparamere slightly projected beyond hind margin of epiphallus, its basal portion rather thickly elongated, but its distal portion sharply pointed and abruptly incurved at apex.
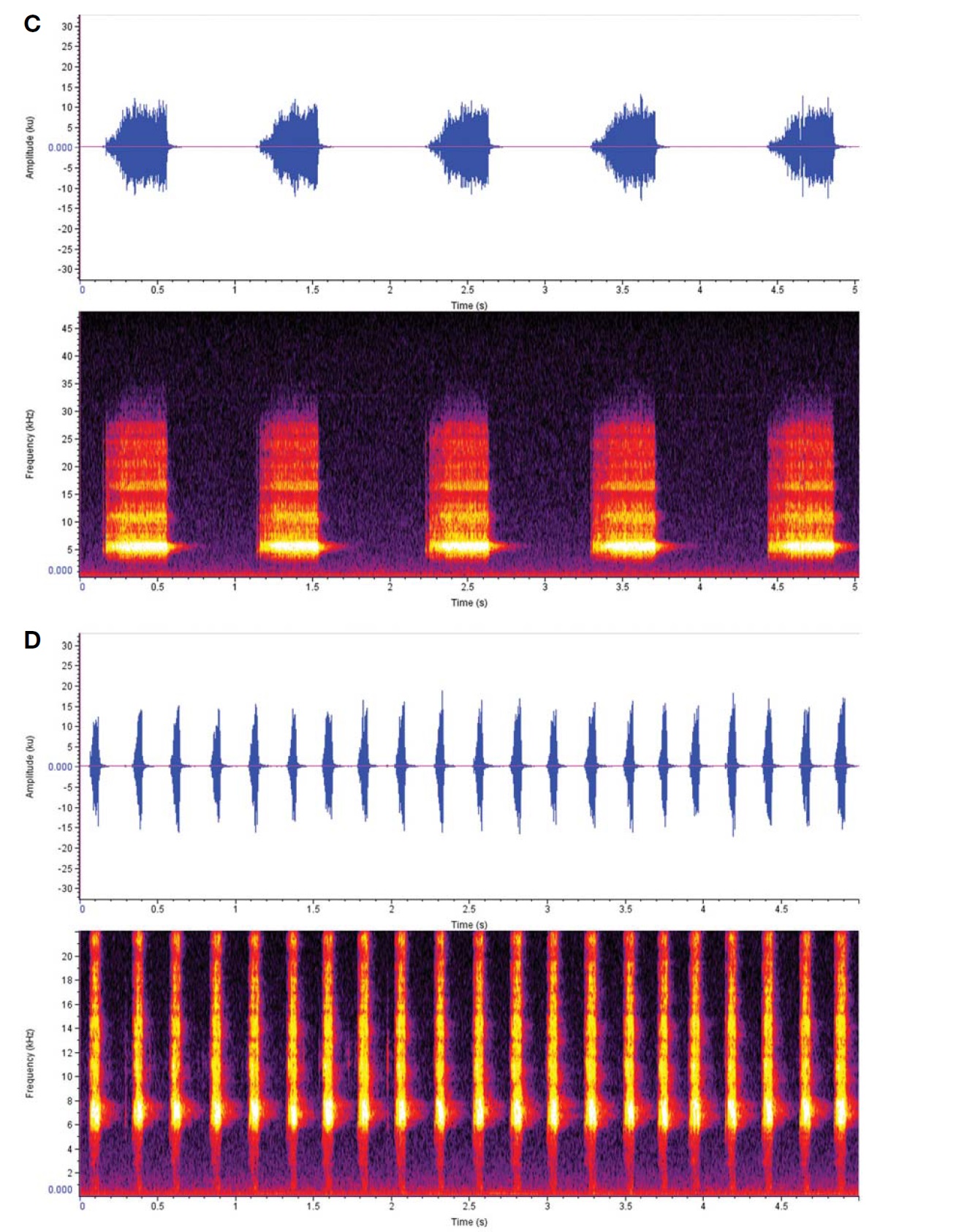
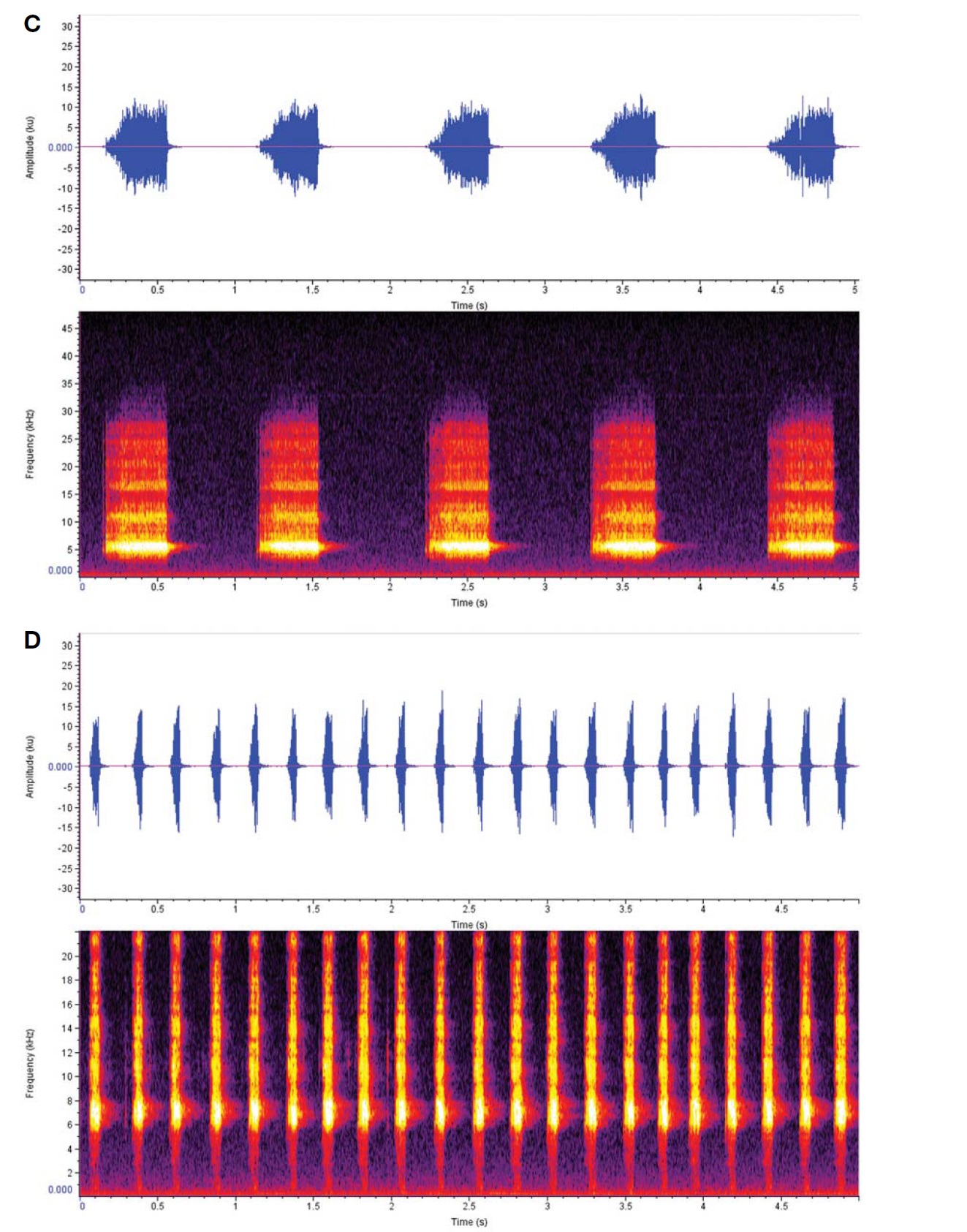
Calling sound (Fig. 5C): Slowly and regularly repeated song phrase; one chirp for one second, but their intervals sometimes irregularly ceased, depending on outer conditions; a chirp lasted 0.45-0.5 s with 46-54 dense syllables, whereas interlude between the chirps taken 0.5-0.55 s at 28° condition; dominant frequency spectrum 4.3-6.2 KHz.
Female (Fig. 1F): Head size similar to male, but tegmina shorter than male, just covering less than half of dorsal abdomen, its apex roundly truncated. Subgenital plate convex, roundly trapezoidal, its hind margin narrowly truncated. Ovipositor as long as hind femora.
Distribution. Korea (new record), Japan, China, Taiwan.
Remarks. This burrowing cricket overwinters as nymphal stages. Calling sounds of the adult can be heard seasonally more earlier than the other
Genus Lepidogryllus Otte and Alexander, 1983
Type species: Gryllus parvulus Walker, 1869 (type locality: South Australia).
Diagnosis. Body coloration dark brown, legs brighter than body, mottled with speckles. Sexual dimorphism not more remarkable than
1*Lepidogryllus siamensis (Chopard, 1961) com. & stat. nov. (Tables 1, 2, Figs. 1G, H, 2D, 3D, 4D, 5D)
Modicogryllus siamensis Chopard, 1961: 280, Pl. 7 (type locality: “Siam”=Thailand); Paik et al., 2010: 77; Paek et al., 2010: 34; Kim, 2011: 49.
Scapsipedus parvus nec Chopard, 1928: 18 (type locality: India); Scapsipedus parves [!]: Choo and Choi, 1983: 49; Velarifictorus parvus: Kwon and Huh, 1994: 50; Kwon et al., 1996: 104.
Material examined (58 specimens). Korea: Chungcheongbuk- do: 1♀ (light trap), Danyang-si, 14-15 Jun 1993, Anonym; Gyeongsangbuk-do: 1♀, Uljin-gun, Sokwang-ri, 1 Aug 1999, Ku DS; Gyeongsangnam-do: 1♀, Sacheon-si, Chukdong- myeon, Baechun-ri, 13 Sep 2001, Paik YH; 1♂ (light trap), Sacheon-si, Waryong-dong, Mt. Waryongsan, Cheongryongsa 11-12 Aug 1999, Sohn JH; 1♂1♀, Jinju-si, Yugok- dong, Nabul-ri, 16 Aug 2012, Kim TW, Kim OS; 3♂ 1♀ (light trap), Jinju-si, Jinseong-myeon, Gajin-ri, Mt. Wolyasan, 13-14 Aug 1999, Park JS et al.; 1♀ (light trap), Goseong- gun, Sanri-myeon, Dongsan-ri, Hyangnobong, 17-18 Aug 1998, Park JS; 2♀ (light trap), Goseong-gun, Sangrimyeon, Osan-ri, Mt. Odusan, 19-20 Aug 1998, Park JS; 2♀, Goseong-gun, Hail-myeon, Deokho-ri, Seoksan, 20-21 Aug 1998, Jeon JS, Kim SY; 4♀ (light trap), Goseong-gun, Hailmyeon, Yongtae-ri, Mt. Sanseongsan, 18-19 Aug 1998, Park JS; 2♂2♀ (light trap), Masan-si, Happo-gu, Jinjeon-myeon, Yeoyang-ri, Osilgol, Mt. Yeohangsan, 11-12 Sep 1999, Park JS; 2♀, Namhae-gun, Bonghwa-ri, Hwacheoncheon, 27-28 Aug 1998, An DH, Sohn JH; 2♂2♀ (light trap), Hapcheongun, Gaya-myeon, Jukjeon-ri, Mt. Gayasan, 19-20 Aug 1999, Park JS; 1♀ (light trap), Hadong-gun, Geumseo-myeon, Galsa-ri, 6-7 Aug 1997, Jeon JS; 1♀ (light trap), Hapcheongun, Bongsan-myeon, Bonggye-ri, Mt. Mangilsan, 22-23 Aug 1998, An TH; 1♀ (light trap), Jinyang, 26-30 Aug 1992, Anonym; 2♂, Geoje-si, Yeoncho-myeon, Songjeong-ri, 11 Jun 2009, Kim TW; Jeollabuk-do: 1♀ (light trap), Muju-gun, 21-25 Aug 1992, Anonym; 1♀ (light trap), Buan-gun, Byeonsanbando, 25 Jun 1991, Ku HJ; Jeollanam-do: 2♂, Suncheon-si, Pungdeok-dong, Dongcheon, 3 Sep 2005, Kim TW; 1♀, Gwangyang-si, Mt. Baekwunsan, 12 Aug 1994, Choi HS; 1♀, ibid. 5 Aug 1990, An SB; 1♀, Gwangju-si, Mt. Mudeungsan, 21 May 1998, Oh KO; 1♀, Hampyeonggun, Gigak-ri, 2 Sep 2000, Park WS; 1♀ (light trap), Yeongam- gun, 21-25 Jun 1991, Anonym; 1♀, Mokpo-si, Mt. Yudalsan, 14 Aug 1992, Oh SH; 1♂3♀ (light trap), Yeonggwang- gun, Isl. Anmado, 19 Aug 1989, Kim JI, Lee OJ; 5♂, Wando-gun, Wando-eup, 1 Jul 2010, Kim TW; 2♂, ibid. 15 Jun 2011, Sung KS; Jeju-do: 2♂, Seoguipo-si, Dongheungdong, Jeongbang-pokpo, 9 Jul 2002, Kim TW; 1♀, Namjejugun, Andeok-myeon, Gamsan-ri, Andeokgyegok, 13 Aug 2004, Kim JK, Kim TW.
Description. Male (Fig. 1G): Head shining black; frons vertical; occiput with six longitudinal pale strips posteriorly; a bright strip transverse between lateral ocelli; interocular distance three times wider than horizontal diameter of eye in dorsal view; narrowest distance between antennal sockets 1.5 times wider than horizontal diameter of antennal socket; median ocellus transversely elliptical, located in dorsum of fastigium of vertex with a pale longitudinal line linked below to clypeal suture; clypeal suture transverse three fifths of head, weakly upraised in mid; scape as long as wide; palpi bright. Pronotum as long as head, two times wider than long in dorsal view; anterior and posterior margins pilose, truncated; dorsal surface brownish, mottled; lateral lobe of pronotum a little deeper than pronotal length, anterior lower margin more bright. Tegmina (Fig. 3D) as wide as hind margin of pronotum, hardly reaching abdominal end; harp veins two or three; chordal veins three; mirror area divided, 1.2 times longer than wide; apical area well netted, 1.2 times longer than mirror area; lateral field a little deeper than lateral lobe of pronotum, with 5-6 longitudinal veins. Fore tibia with largely oval-shaped outer tympanum, its length three times longer than wide; inner tympanum small and rounded, 1.2 times longer than wide. Hind femur 2.7 times longer than middle femur. Hind tibia with 5-6 pairs of large dorsal spines on both sides, the largest inner apical spur as long as a half of basitarsus. Hind basitarsus 1.7 times longer than remaining two tarsomeres combined length; 5-7 dorsal spinules on both sides; inner apical spur as long as a half of last tarsomere. Cercus as long as two thirds of hind femur. Subgenital plate shortly conical, its hind margin truncated, deeply notched in mid.
Genitalia (Fig. 4D): Epiphallus with narrow central bridge, its width wider than the length of lateral lobes; median lobe absent; inner and hind margin with a lot of hairs. Ectoparamere slightly surpassing lateral lobe of epiphallus, divided into two triangular projections at apex, which are almost subequal in width, strongly and shortly incurved forming forked terminal.
Calling sound (Fig. 5D): Short and quick song phrase; buzzy chirps regularly repeated 4-5 times for one second; a chirp with undulating 11-12 syllables at 28℃ condition; dominant frequency spectrum 5.8-7.3 KHz.
Female (Fig. 1H): Head with more round frons, mouth parts shorter than in male; interocular distance four times wider than horizontal diameter of an eye. Cercus slightly shorter than ovipositor. Subegenital plate roundly triangular, its hind margin weakly notched. Ovipositor straight needle-shaped, as long as hind femur; apex sharply pointed, subapically dilated, without serration.
Distribution. Korea, Japan, Taiwan, Thailand, India (Oriental Region); Hawaii (introduced). China possibly included in this wide distributional ranges of this cricket, pursuing after records of
Remarks. This cricket overwinters as nymphal stages in southern localities. Calling sounds of adult can be heard from May. Their sounds mixed with the ones of
Key to the Species of Velarifictorus and Lepidogryllus from Korea
1. Fore tibia with large outer and small inner tympana. Male tegmina narrower and elongate, its apical area distinctly longer than mirror area; female tegmina as long male tegmina ?????????????????????????????????????? Lepidogryllus siamensis
- Fore tibia with only outer tympanum. Male tegmina wider and relatively short, its apical area shorter than or as long as mirror area; female tegmina distinctly shorter than male tegmina ??????????????????????????????????????????????????????????????????? 2
2. Male with elongate mandibles, longer than upper half of head, thus clypeal suture transverse upper than mid in frontal view; ovipositor shorter than hind femur ?????????????????????????????????????????????????????? Velarifictorus aspersus borealis
- Male with moderate or normal mandibles, shorter than upper half of head, thus clypeal suture transverse lower than mid in frontal view; ovipositor longer than hind femur ???????????????????????????????????????????????????????????????????????????????? 3
3. Head with bright ∧-shaped strip running transversely between lateral ocelli ?????????????????????? Velarifictorus micado
- Head without distinct transverse strip between lateral ocelli, that reduced to ecdysial feeble line ?????????????????????????????????????????????????????????????????????????????? Velarifictorus ornatus
Korean name: 1*탈귀뚜라미
Korean name: 1*극동귀뚜라미
Korean name: 1*봄여름귀뚜라미(신칭)
Korean name: 1*샴귀뚜라미(신칭)