Tropical tasar culture involves rearing of tropical tasar silkworm
Experiments of present work were undertaken at Silkworm Breeding and Genetics laboratories and rearing of silkworms at the farm of Central Tasar Research & Training Institute, Ranchi during the second crop rearing season (September-November, 2012).
>
Grainage and raising of F1 hybrid
Grainage was conducted during September 2012, Daba cocoons harvested during the first crop (July-August 2012) were collected from the Germplasm bank, Silkworm breeding section, Central Tasar Research &Training Institute. Laria cocoons were collected from forest area of Peterbar, Jharkhand. The good cocoons were sorted based on the sex. A total of 100 male and identical numbers of female cocoons each of two ecoraces were selected and stored at normal grainage conditions until the emergence of moth. When emergence started, crossings were made as Laria x Daba, besides the parental stocks. Mated female moths of all the batches were kept for oviposition for 3 days at normal grainage conditions.
The fecundity was calculated as total number of eggs laid by a single mother moth in three days of egg laying (Sinha, 1998). Hatching rate was recorded (Saheb
>
Preparation of samples for biochemical studies
Hemolymph, fat body and midgut of Daba, F1 hybrid and Laria of both the sexes were collected separately during 5th day of V instar for the biochemical studies which is known to be the most active stage of tasar silkworm in terms of metabolism. Hemolymph and fat body samples were collected by dissecting out the larva and further processed (Lokesh
The total protein concentration was assayed as described by Lowry
The date generated in the present study is the average values of three replicates (n = 5 for biochemical analysis and n = 100 for quantitative trait analysis). One-way ANOVA was carried out using SPSS 10.0 statistical package to test the significance
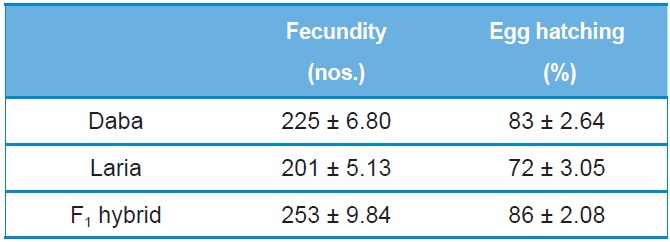
[Table 1.] Fecundity and egg hatching of Daba, Laria and F1 hybrid (Mean ±SE)
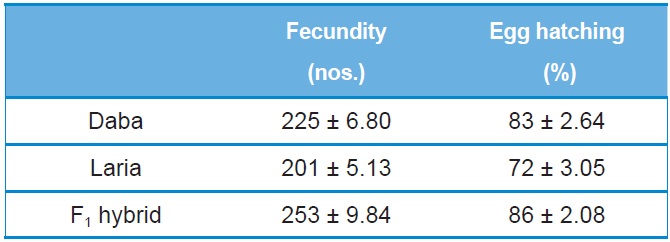
Fecundity and egg hatching of Daba, Laria and F1 hybrid (Mean ±SE)
of differences, between the means of individuals and among the groups studied. Comparisons were performed with Duncan’s Multiple Range Test (DMRT, P<0.05) (Duncan, 1955).
The performance of the F1 hybrid (Laria x Daba) tasar silkworm was analyzed and compared with its parental lines for fecundity, egg hatching and biochemical parameters are recorded.
The performance of F1 hybrid was recorded with significant improvement in the fecundity (
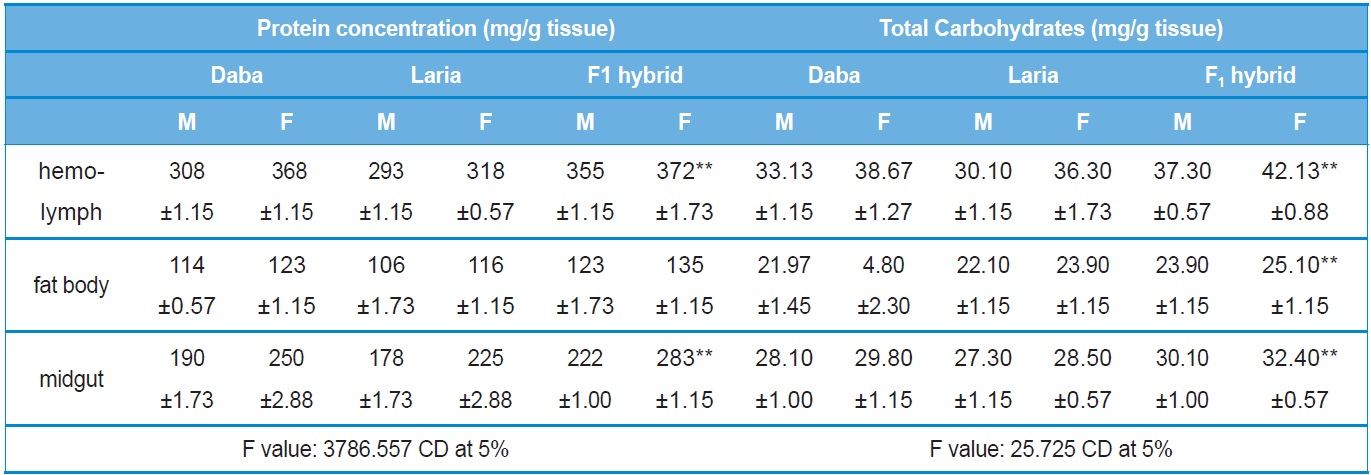
Protein concentration was recorded highly variable between the different batches (Daba, Laria and F1 hybrid) and the tissues studied. Higher protein concentration was recorded in the hemolymph of F1 hybrid female larvae (372.00±1.73 mg/ml) followed by Daba female larval hemolymph (368.00±1.15 mg/ml) and the lowest concentration (106.00±1.73 mg/ml) was recorded in the fat body of male Laria larva. A significant increase in the protein concentration (
The level of carbohydrates was recorded the significantly higher (
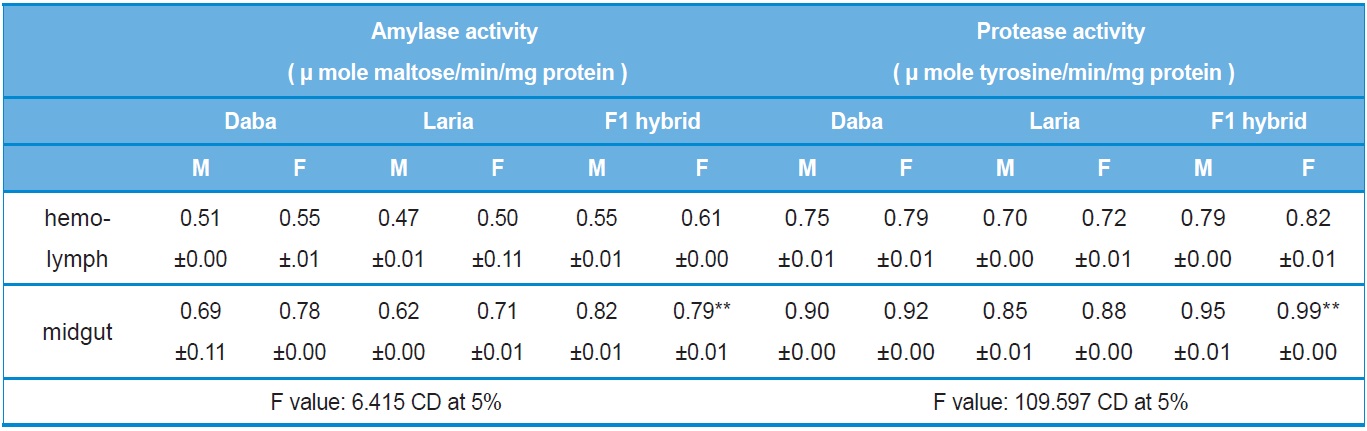
The level of activity showed significant variations among the larval batches viz., Daba, Laria and F1 hybrid. Significantly higher (
>
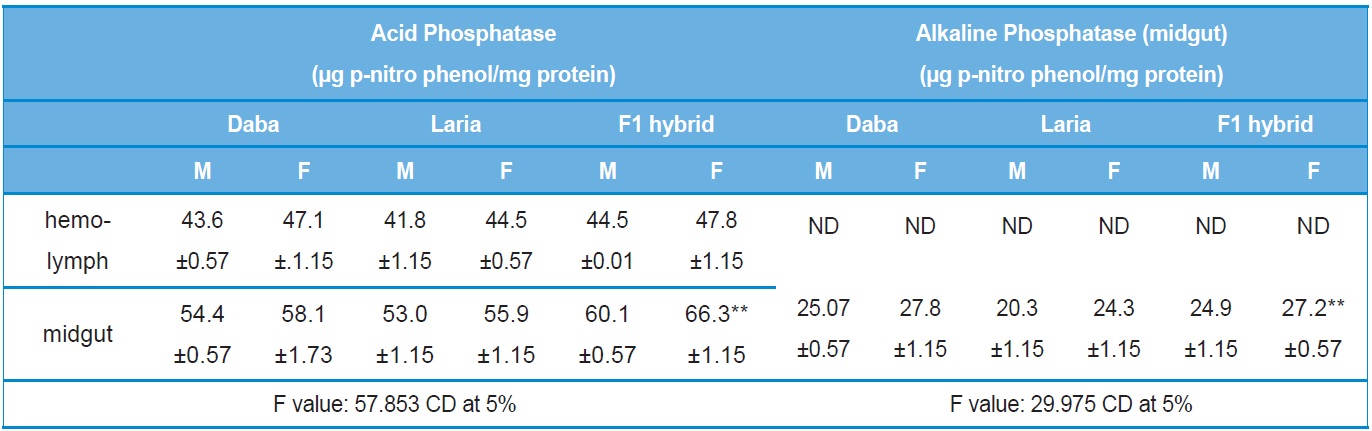
Acid and Alkaline phosphatase
An increase in the phosphatase activity was found in the midgut tissue of F1 hybrid larva the level of the activity was significantly higher to the parental lines (Table 4).
>
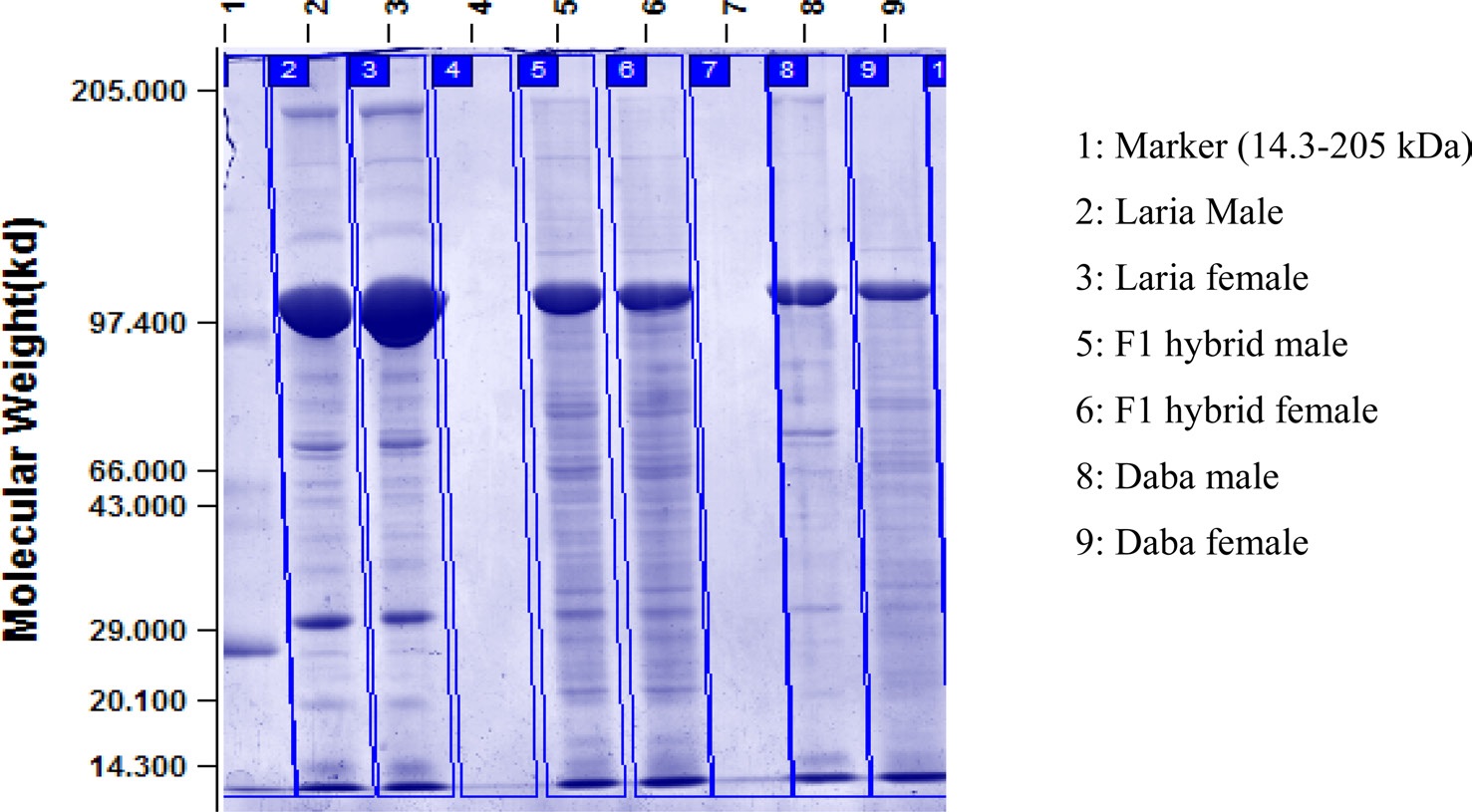
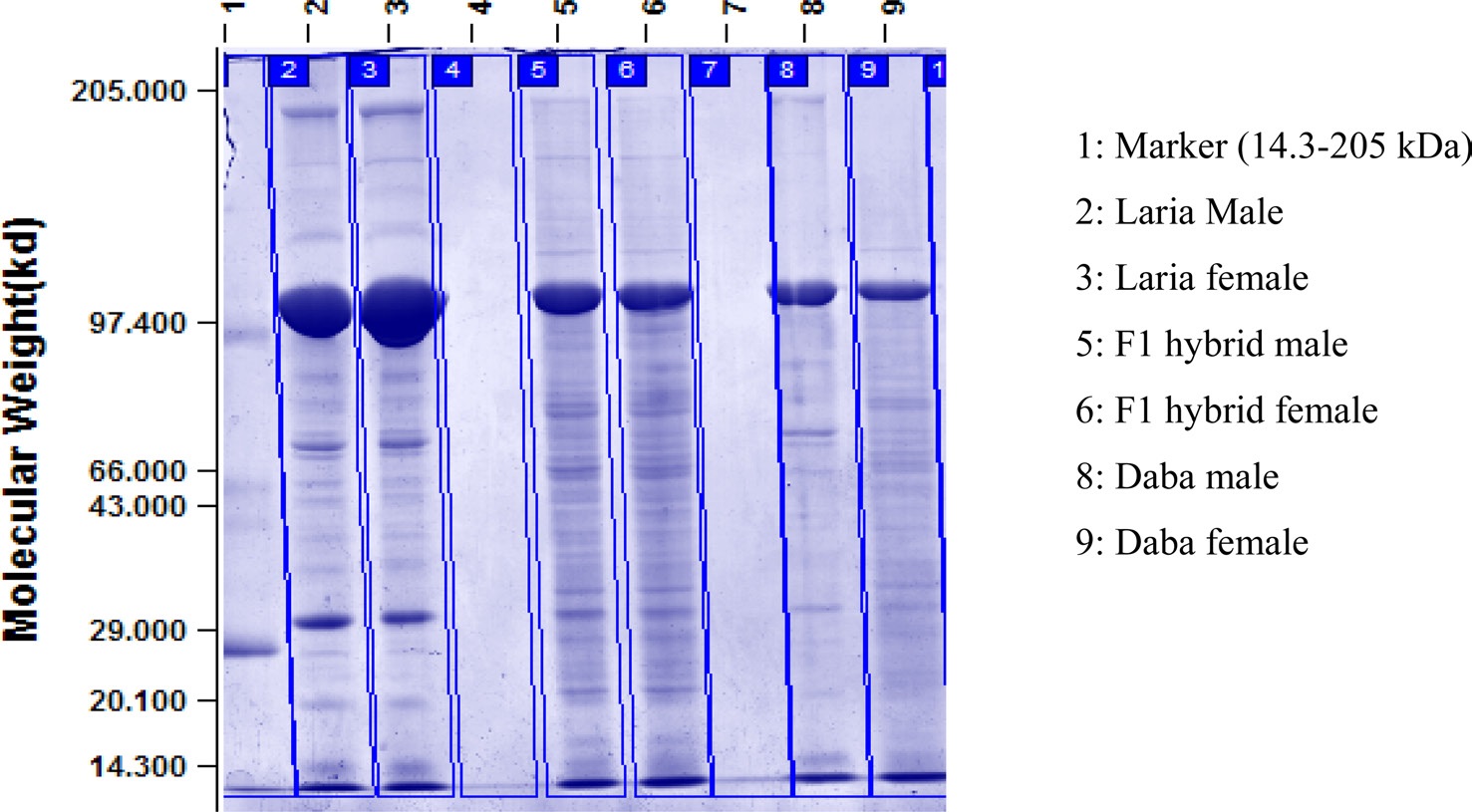
Electrophoresis of hemolymph protein
Qualitative analysis of haemolymph proteins in the V instar larva of Daba, Laria and F1 hybrid of tasar silkworm was recorded (Fig. 1) Polymorphic variations were noticed with regard to the number of protein bands in male and female also in different batches viz., Daba, Laria and F1 hybrid. Protein bands of 98 KDa, 70 kDa, 45 kDa,
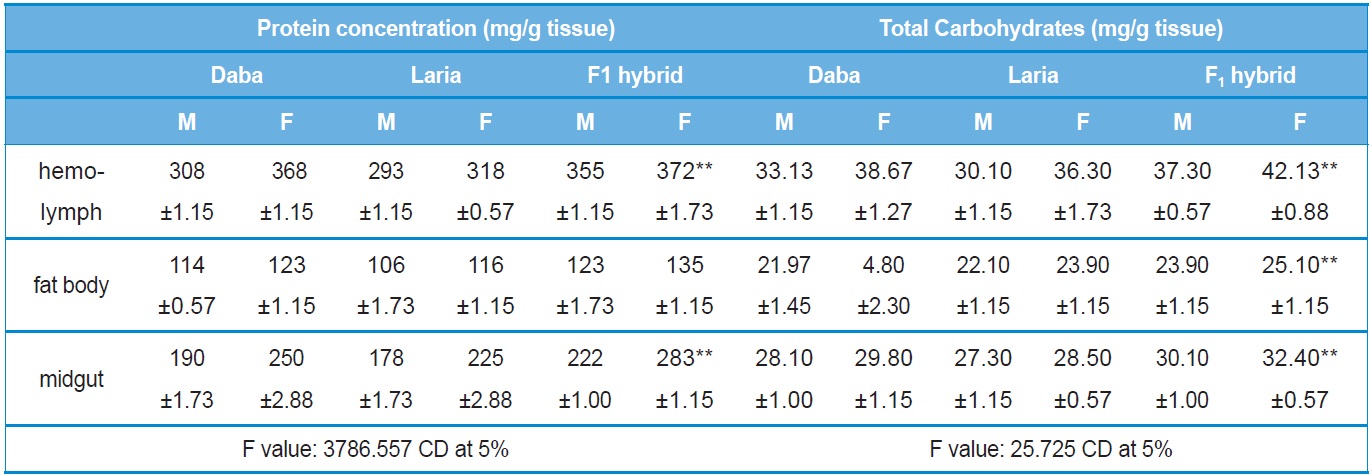
Quantitative analysis of total protein and carbohydrate concentrations in the different tissues of Daba, Laria & F1 hybrid (Mean ±SE)
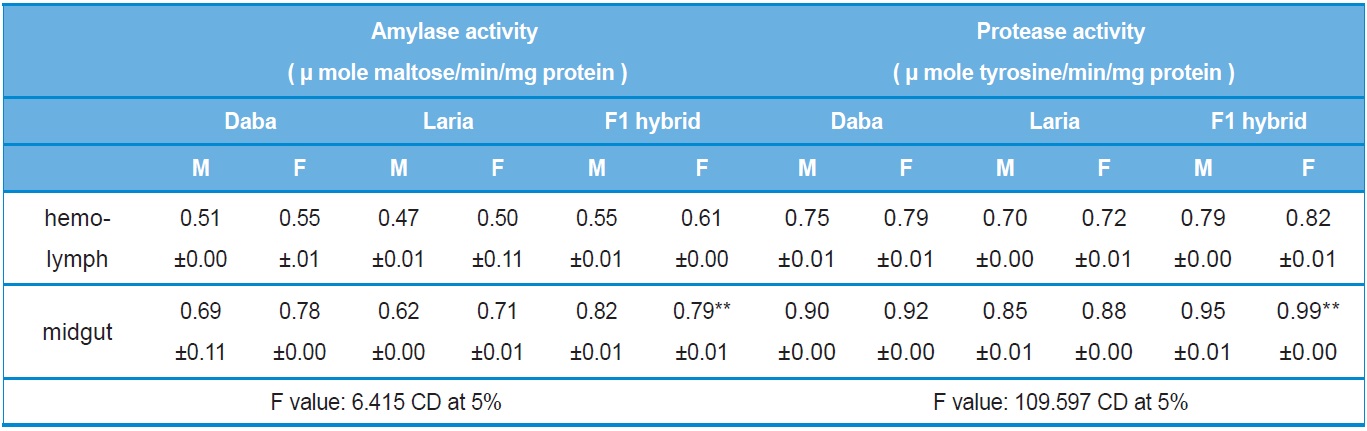
Activity analysis of digestive enzymes in the different tissues of Daba, Laria & F1 hybrid (Mean ±SE)
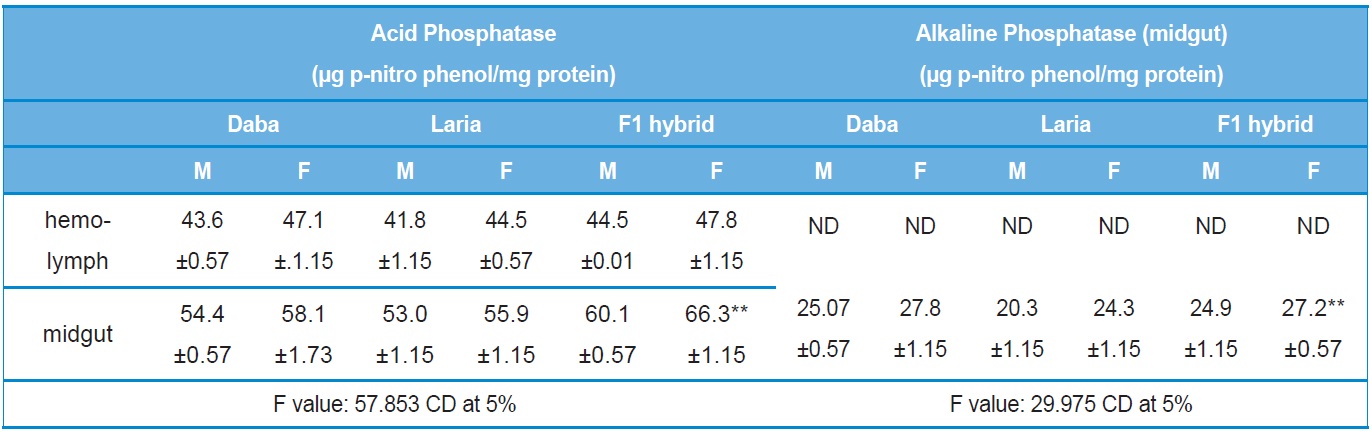
Activity analysis of Acid and Alkaline Phosphatase in the different tissues of Daba, Laria & F1 hybrid (Mean ± SE)
28 kDa and 14 kDa are found common in all the hemolymph samples. Additional number of protein bands was recordedd in the hemolymph sample of F1 hybrid ranging between 29 kDa to 66 kDa compared to the samples of Daba and Laria. Conspicuously stained bands of 67 kDa appeared in the hemolymph samples of F1 hybrid.
The quantitative characters in silkworm are highly variable and have a greater economic value. F1 hybrids are most commonly used for commercial cocoon production because they represent high heterosis for most of the economic characters (Singh
Higher concentration of the total protein and carbohydrates recorded in the present study for F1 hybrid is attributed to the fact that these larvae have acquired robustness due to the interecoracial crossing and fed higher quantity and quality leaves in turn efficiently converted the dietary food during its feeding stage, the corresponding level of proteins were maintained in the hemolymph, midgut and fat body. Also, many authors have similarly recorded higher proteins in the hemolymph of female earlier (Babu
Study of protein banding patterns by SDS-PAGE in the hemolymph samples drawn from different larval batches revealed that the number of protein bands and specificity of bands vary between the larval batches and also between male and female. The presence and absence of few protein bands at different molecular weight indicates the synthesis of a particular protein to meet the physiological need. Protein bands of 98 kDa, 70 kDa, 45 kDa, 28kDa and 14 kDa are found common in all the hemolymph samples of all the larva batches. This could be of species specific proteins, presumed to found in all the ecoraces of the
Better performance of F hybrid of Laria x Daba in almost all the parameters studied in the present study indicates the possible influence of hybridization. Also a positive heterosis effect in the F1 hybrid this envisages the possible utilization of F1 hybrids from the crosses of wild and semi-domesticated ecoraces of tropical tasar silkworm for optimal silk productivity. However, biochemical analysis envisages that, there will be possibilities of breeding with specific parents obtaining desirable commercial traits (silk yield and fecundity) are an impending tool to optimize heterosis for genetic improvement in the desired direction. Moreover, these biochemical parameters can be used as markers to analyze the genetic improvement in the tasar silkworms especially in hybrids.