At the regional scale, differences in climate, physical geomorphology, biogeographic history, and large-scale patterns of human land use can lead to differences in available species pools (Rickelfs 1987, Zobel 1992, Foster et al. 2003). Within the region, surrounding land uses, site isolation, and site area influence plant species composition by determining rates of propagule input (Matthews et al. 2009). Wetlands are also affected of hydrology, physico-chemical environment (Mitsch and Gosselink 2007). Hydrology regime is considered to be importance in determining characteristics of wetland plant communities (Keddy 2000, Mitsch and Gosselink 2007). Depth, duration, and frequency of flooding are major controls of seed germination and establishment (van der Valk 1981, Casanova and Brock 2000), boundaries between woody and herbaceous vegetation (Toner and Keddy 1997), plant productivity and diversity (Fennessy et al. 1994), and species composition (Weiher and Keddy 1995). It also determines the frequency and intensity of flood disturbance, which creates topographic differences and erosion (Naiman and Decamps 1997). Apart from those, other important factors of the plant composition in wetlands include fertility, salinity, competitiveness between plants and herbivores (Keddy 2000). Soil fertility is also a determinant of plant community and diversity in wetlands (Weiher and Keddy 1995) because wetland soils are both the medium in which many of the wetland chemical transformations take place and the primary storage of available chemicals for most wetland plants (Mitsch and Gosselink 2007), so that nutrient enrichment could increase growth of invasive species (Green and Galatowitsch 2002, Kercher and Zedler 2004, Rickey and Anderson 2004, Rejmankova 2011) and these environmental conditions could lead a different stage of succession.
As paddy cropping was especially important economic activity in Asia, many paddy fields and paddy terraces in mountainous valleys were developed for intensive agricultural systems (Kim et al. 2006, Park et al. 2006). Since the improvement of socio-economic conditions, there was low economic benefit due to decrease in consumption of rice, rural exodus of farmers, and government agricultural policies aiming to reduce overpopulation, paddy fields have been increasingly abandoned (Park et al. 2006, Byun et al. 2008). Especially, because of being located at mountainous valley, paddy terraces were inaccessible and unfavorable farming conditions such as inclining topography, small terraced plots, and levee grasses that were hard to mow, they have been getting non-cropped land and they did not be cared (Fukamachi et al. 2005). Although they were the largest man-made wetland ecosystems at first, they have changed into natural wetlands after abandonment (Lee et al. 2002, Kim et al. 2006, Yamada et al. 2007). Therefore, they could provide a function of foods and protection of fowl, and habitats which could increase biodiversity (Comin et al. 2001).
Until now, abandoned paddy fields have been studied for plant succession within near-regional but these abandoned paddy terraces were distributed to several locations with mountain ranges in Korea. After abandonment, they were transformed from artificial wetlands into natural wetland by being affected by environmental condition. Diverse plant species could be established according to hydrology regime and soil condition and they could be in the different development stage of plant community. Here, we investigated physico-chemical characteristics and plant community of abandoned paddy ter-races
which represent three regions disconnected by the mountain ranges, which divide watersheds and climate zones. The object of this study is to reveal the relationship between characteristics of environmental conditions and early development of plant community at abandoned paddy terraces in South Korea.
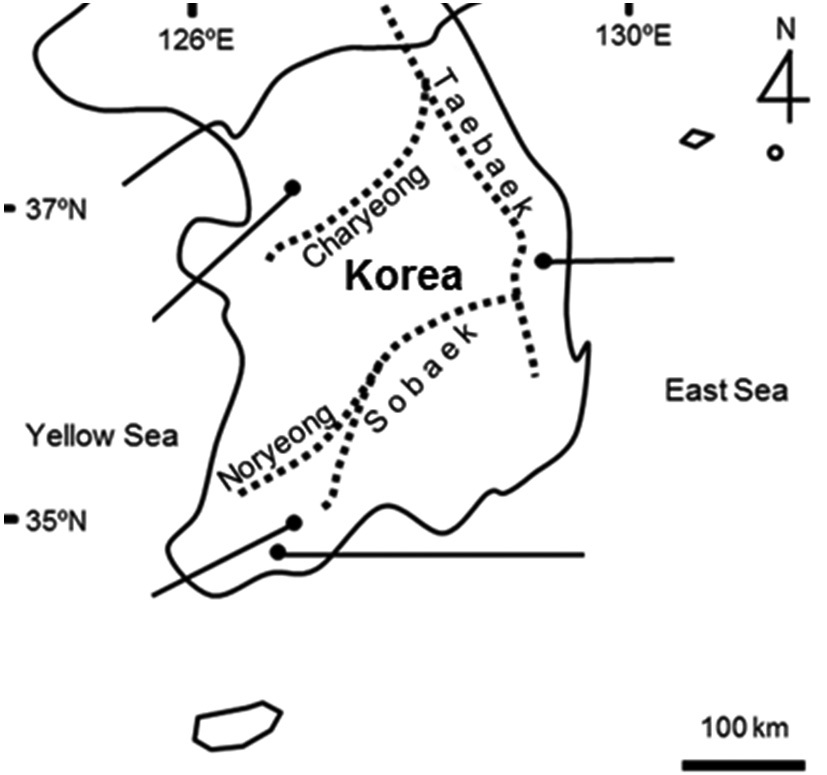
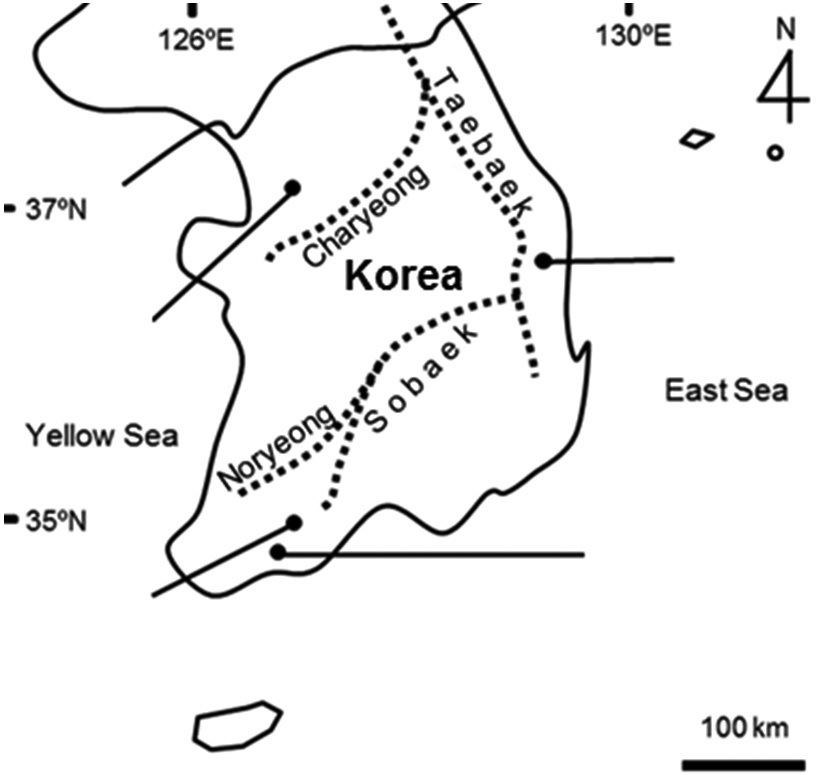
Study sites were located at mountainous valley in Ansan (site A, 37˚17′10.68″N, 137˚55′25.96″E, 107 m a.s.l., 6,353 m2) in Gyeonggi-do, Uljin (site B, 36˚54′26.43″N, 129˚24′46.47″E, 35 m a.s.l., 2,345 m2) which was located 0.33 km off the coast in Gyeongsangbuk-do, Gokseong (site C, 35˚15′53.38N, 127˚16′09.68″E, 115 m a.s.l., 5,102 m2), and Boseong (site D, 34˚48′21.53″N, 127˚04′55.96″E, 158 m a.s.l., 14,020 m2) in Jeollanam-do in Korea (Fig. 1). They represent three regions distributed by the mountain ranges which determine watersheds and climate zones. We selected study sites which were approximately abandoned since between 10 and 15 years, composed of over five floors, and well-maintained in terms of levee structure or water regime after being uncared. Ground matrix of four study sites was granite gneiss (Korea Institute of Geoscience and Mineral Resources 2012). The main water source was surface water. Even though monsoonal climate experiences heavy rain during summer season, the
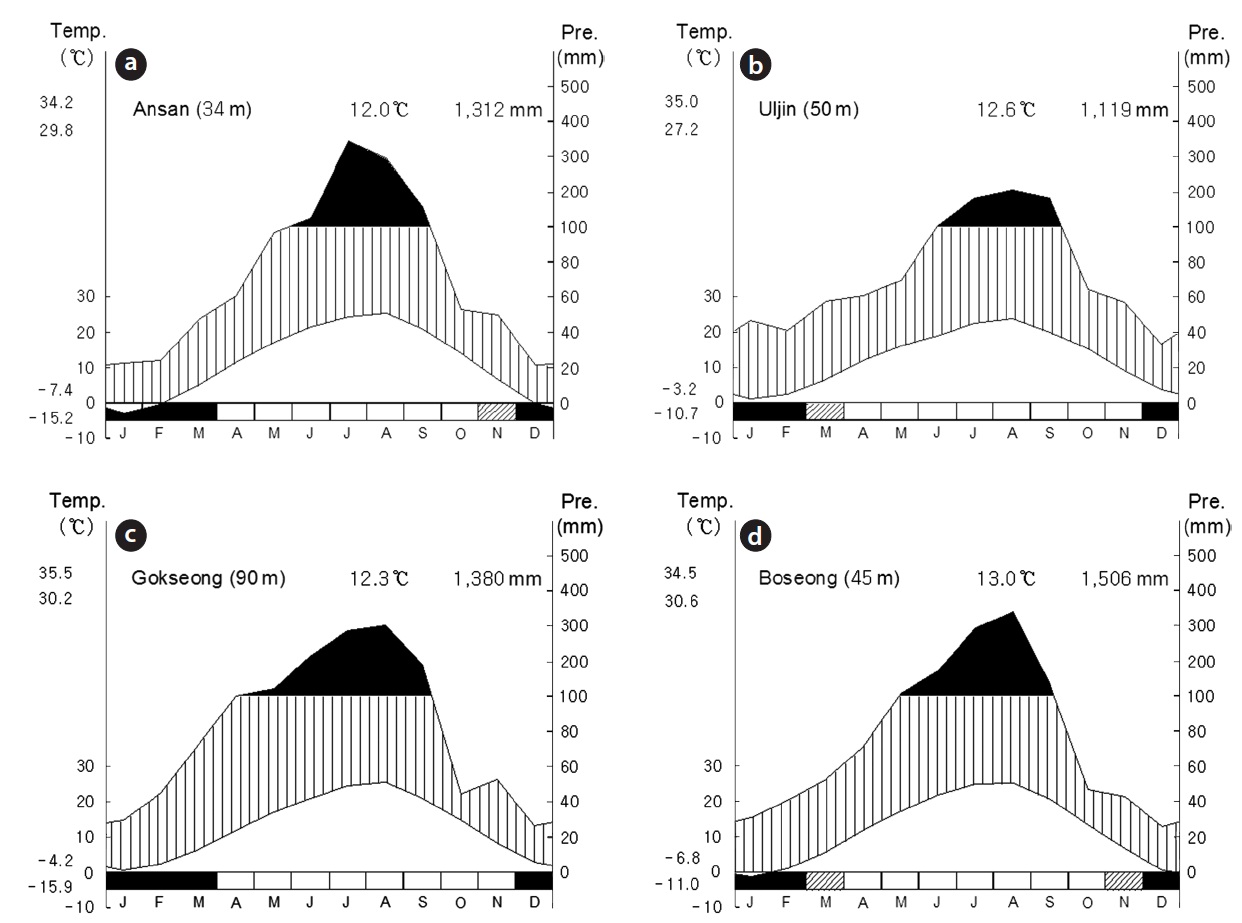
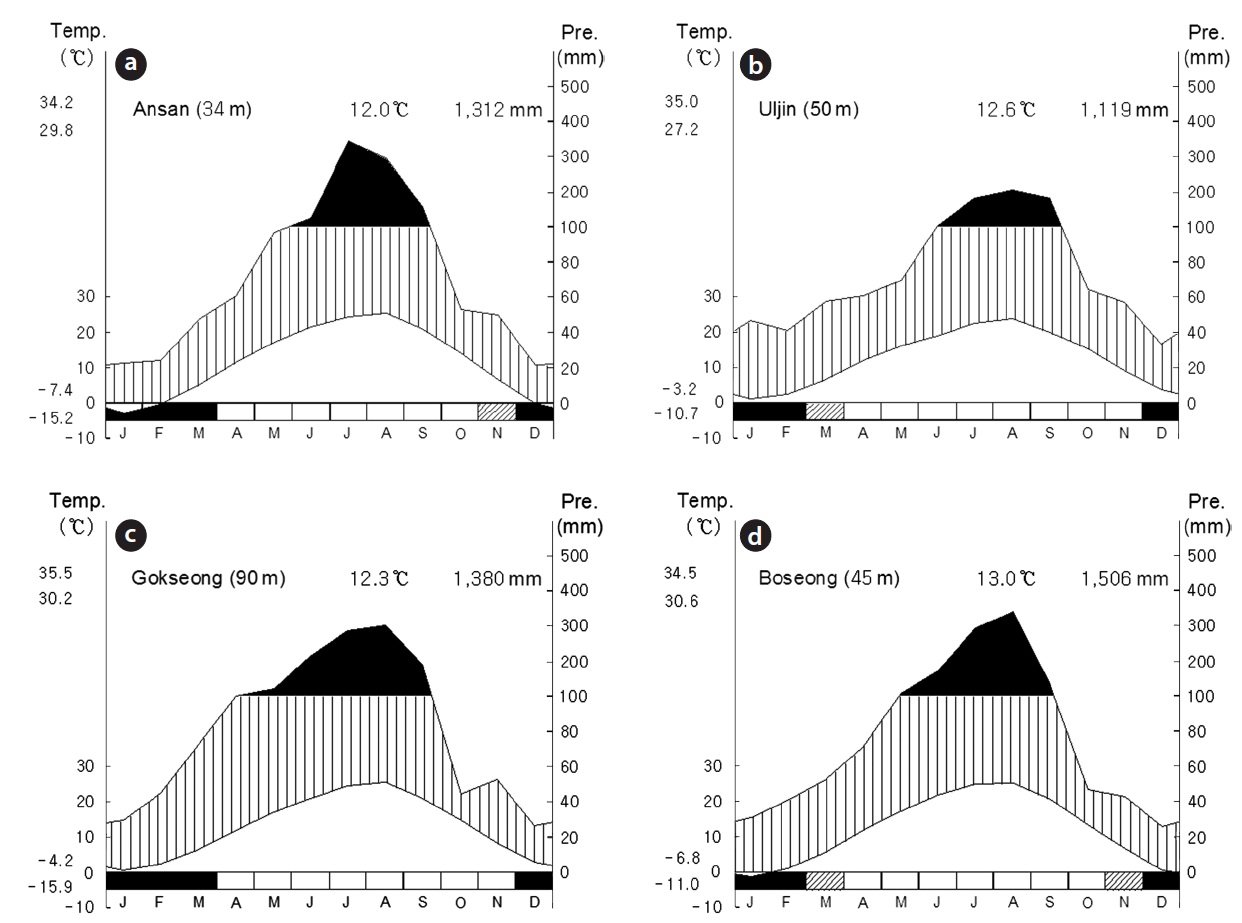
water depth in these sites was kept at a constant level by slopes and dikes with excess water flowing out downhill. Mean temperature and precipitation for 30 years were quietly different among the study sites overall (Fig. 2). Especially, mean precipitation of site B was lower than the other sites. Site C and D located in south showed more amount of mean precipitation and higher mean temperature than the other sites.
>
Soil condition and water depth
Soil samples were randomly collected at five points in each floor with a depth of 0 ~ 5 cm from the surface by using a soil hand auger. Gravel and large organic debris were removed from samples by being passed through 2 mm sieve (standard sieve #10). Soil texture was determined using the hydrometer analysis method and the texture triangle of USDA (Carter 1993). Water content was determined after drying the samples at 105℃ in an oven for overnight (Topp 1993). Organic matter content was analyzed by the loss-on ignition method (LOI) (Boyle 2004). Soil solutions were prepared by mixing the soil samples with distilled water at a mass ratio of 1 to 5 and pH and conductivity (CON) were measured, using a pH meter (AP63; Fisher, Hampton, USA) and a conductivity meter (Model 311; Corning, Lowell, USA), respectively. NO3-N and NH4-N were extracted with 2 M KCl solutions (Kim et al. 2004) and measured colorimetrically using the hydrazine and indophenol methods, respectively (Murphy and Riley 1962, Kamphake et al. 1967). PO4-P was extracted with Bray No. 1 solution (Bray and Kurtz 1945) and measured colorimetrically using the ascorbic acid reduction method (Solorzano 1969). Available K+, Ca2+, Na+, and Mg2+ were extracted with 1 N ammonium acetate solution (Allen et al. 1974) and measured with an atomic absorption spectrometer (Model AA240FS, Varian, Palo Alto, CA, USA).
Water depth was repeatedly measured at five points at each floor. Water depth was measured during the early growing season before the rainy season in 2011.
>
Flora and vegetation composition
The flora list and vegetation map about dominant species were recorded at each floor during summer season in 2011. Each floor in the same area was to be a quadrat and there was repetition at the study site. The flora was investigated by passing all over the floors. Figuring out the dominant species in the map of the vegetation composition was determined by showing over 10% at each floor. The taxonomic nomenclature designated by Lee (2003) was used to identify each plant species. Species richness was obtained by counting the number of species in the whole sites. The Shannon-Weaver diversity index (
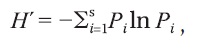
where
To examine and identify the similarities and/or differences in physico-chemical characteristics and plant community, correspondence analysis (CA) was conducted using the vegan-package 2.0-5 (Okasanen et al. 2012) in R program version 2.15.1. The CA ordinations based on the environment factor-site matrix table and the species-site
[Table 1.] Soil conditions and water depth in abandoned paddy terraces (mean ± SD)
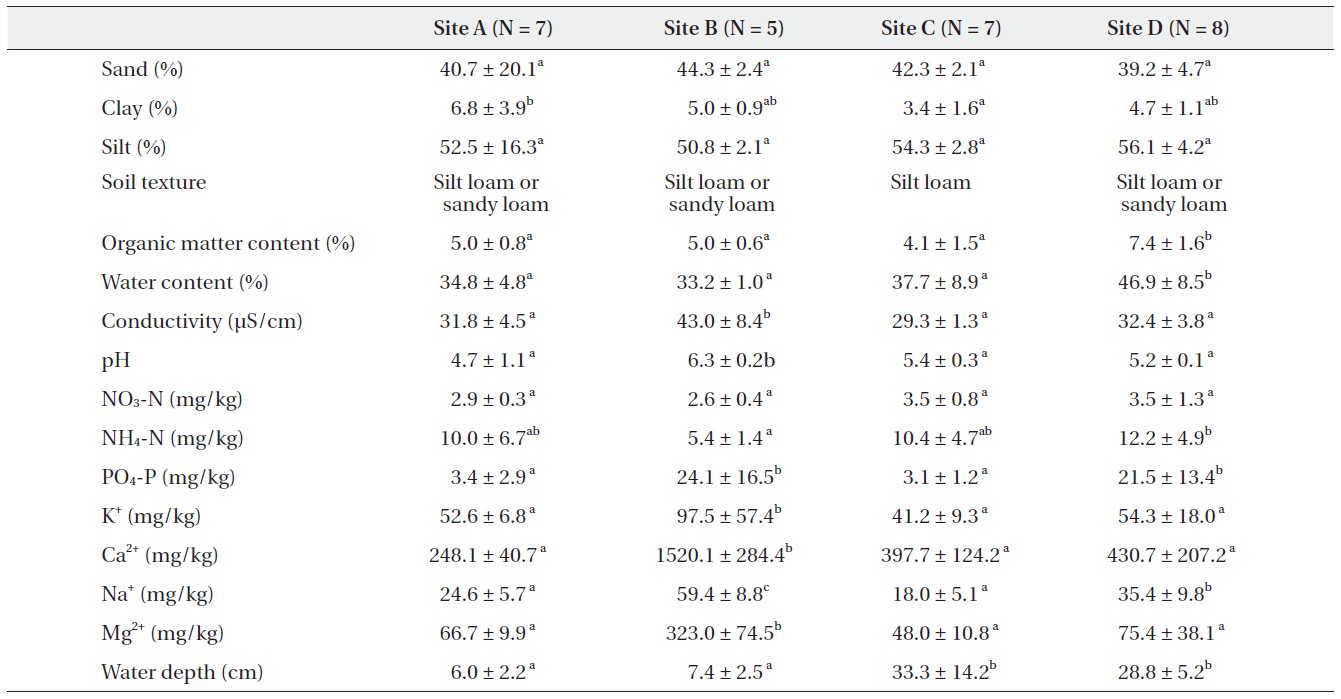
Soil conditions and water depth in abandoned paddy terraces (mean ± SD)
matrix table were carried out. The specific cover data was placed into the species relative coverage.
Duncan’s post-hoc tests were conducted to identify specific differences at the 5% significance level using SPSS version 20.0 for Windows (SPSS, Inc., Chicago, IL, USA).
>
Soil condition and water depth
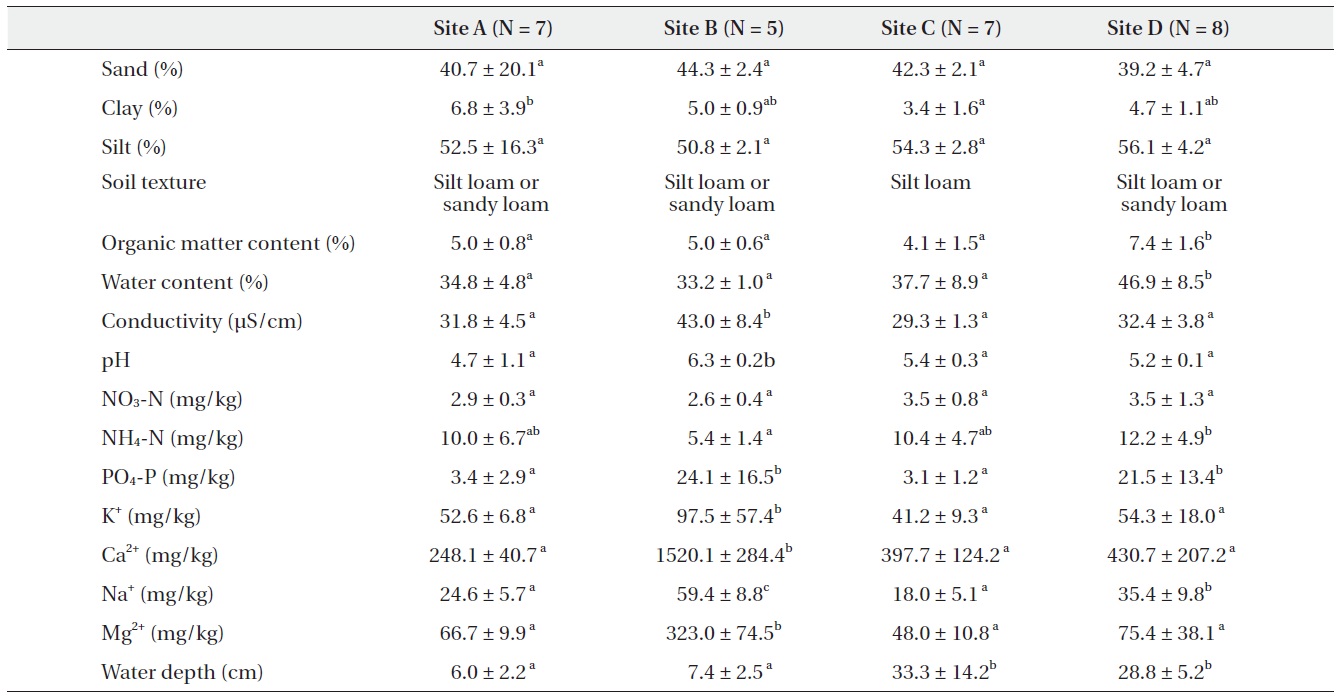
There were significant differences among abandoned paddy terraces in soil characteristics (Table 1). Soil texture was silt loam or sandy loam in most floors. Water contents and organic matter contents within and among abandoned paddy terraces were similar, but site C was slightly higher than the others. The conductivity ranged from 30 to 40 μS/cm in all abandoned paddy terraces and did not show significant difference among abandoned paddy terraces but pH ranged from 4 to 6 and showed a little difference among the abandoned paddy terraces. NO3-N was similar in all abandoned paddy terraces but NH4-N and PO4-P were quite different among abandoned paddy terraces. NH4-N was the least in site B. PO4-P was about eight times higher in site B and D than the others. The contents of major cations were mostly higher in site B which was located near the seashore than the others. Especially, Mg2+
and Ca2+ contents were much higher than the others. The average water depth was higher in site C and D than the others, and site C particularly showed a high gradient of water level among floors.
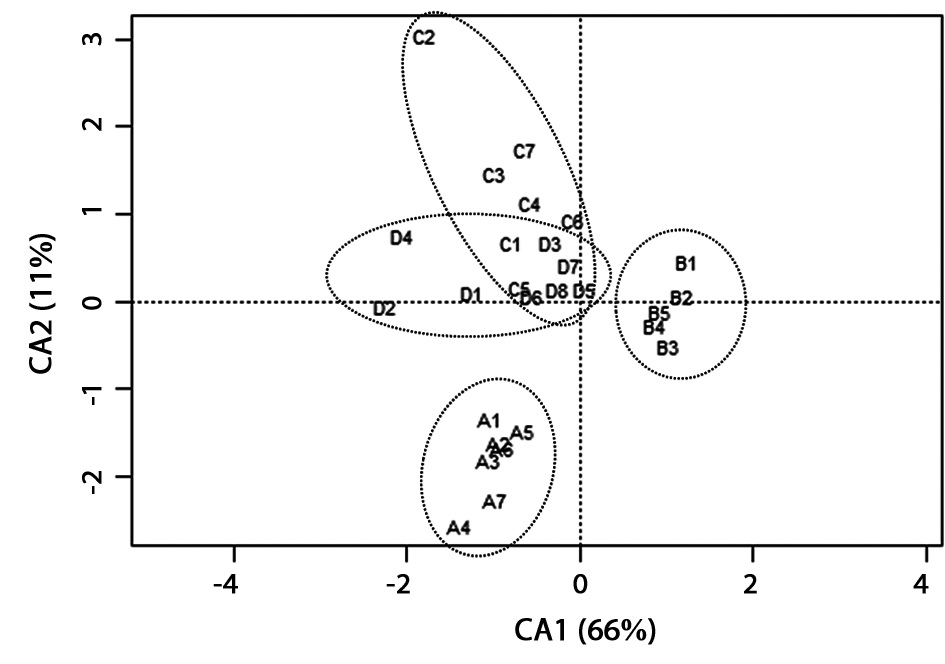
To identify the similarities and/or differences in physico- chemical characteristics among the floors and abandoned paddy terraces, CA ordination was carried out (Fig. 3). In CA ordination, axes 1 and 2 accounted for 66.3% (eigenvalue = 0.10) and 11.5% (eigenvalue = 0.02) of total variance, respectively. Sites with relatively strong loadings on axis 1 and 2 were water depth (score = -0.79 and 0.69, respectively). As a result, floors in the same site were completely grouped but the sites were divided into each location, and site C and D were overlapped in several floors.
>
Flora and vegetation composition
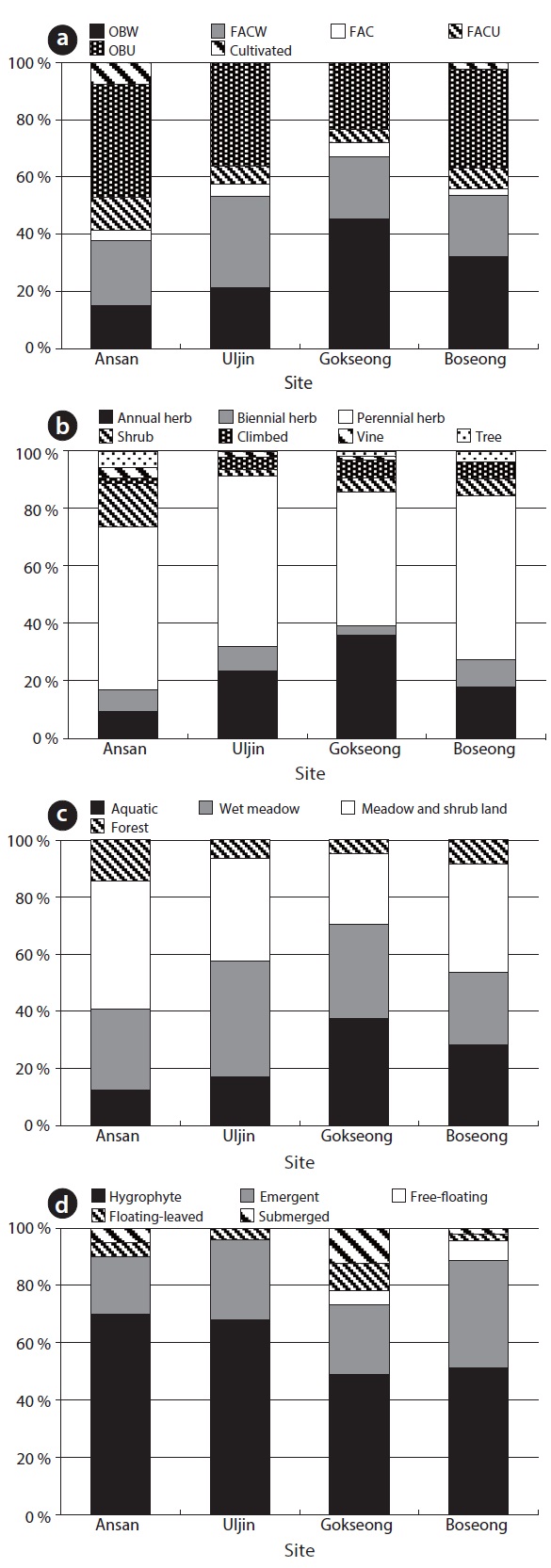
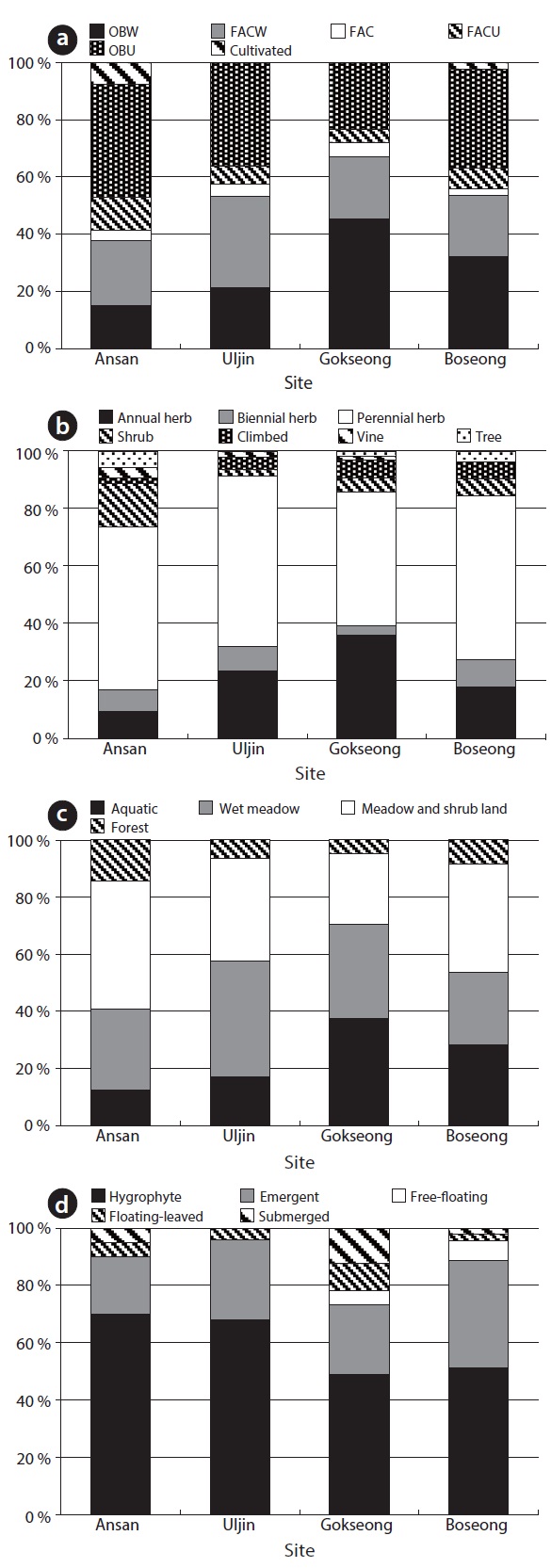
A total of 141 species were recorded in four sites. According to wetlands indicator category (US Fish and Wildlife Service 1996), there was a large amount in obligated wetland plant (OBW) and facultative wetland plant (FACW), but obligated upland plant (OBU) was also counted highly in this study (Fig. 4). Particularly, a large amount of OBW was showed in site C and D which were higher water level than the others, and site C was especially showed the highest ratio of OBW. About life-forms of plants, perennial herbaceous plants largely took up and then annual herbaceous plants were occupied. Site A showed a large coverage of shrub. Site C comparatively showed a high ratio of annual herb because site C showed a gradient of the water depth. Most of the plants were hygrophyte, indicating mostly aquatic environment, wet
meadow or meadow and shrub land. In particular, submerged and floating-leaved plant communities occurred at high ratio in site C. Additionally, rare species
Mean of species richness per floor and diversity index were 22.67 ± 6.25 and 1.56 ± 0.25, respectively (Table 2). Species richness was the largest in site D and the least in site A. However, species richness per square meter was not different between Site A and D and the highest in site B. Shannon’s diversity was the largest in site A and B, but the least C.
Among the 141 species, a total of 93 species occurred
[Table 2.] Species richness and diversity index.

Species richness and diversity index.
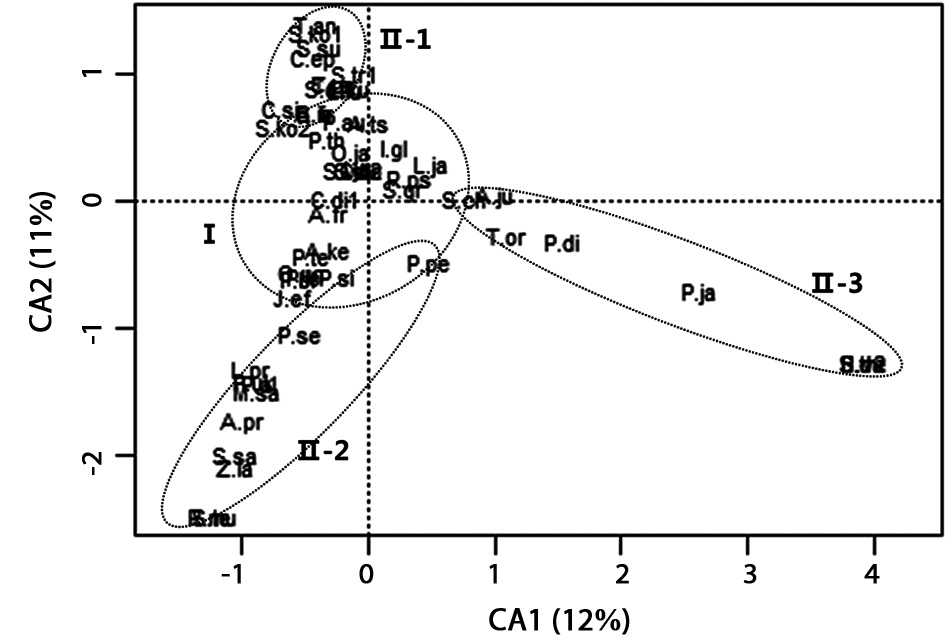
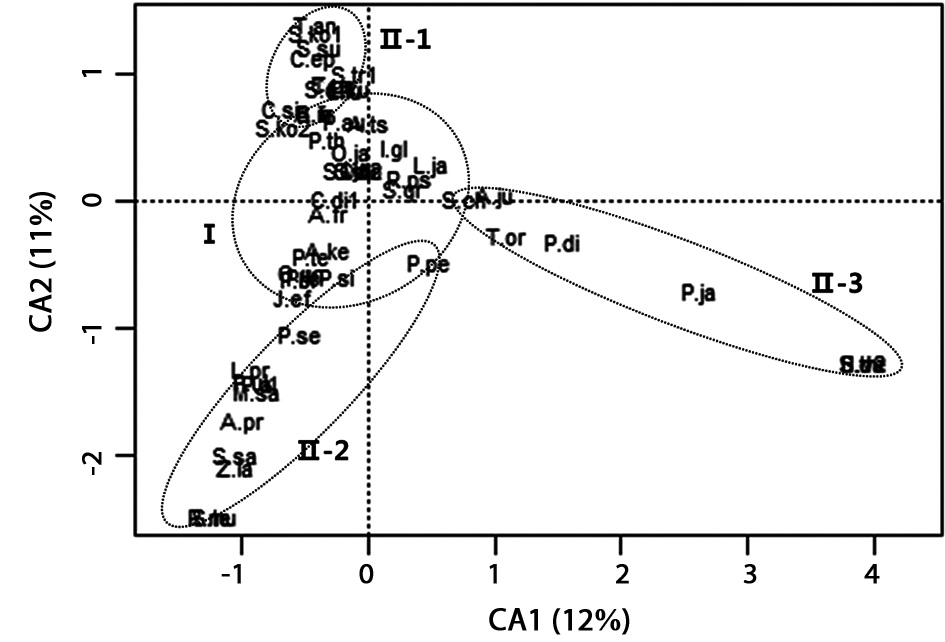
in all sites. A total of 55 species were recorded as dominant species by sum of the whole sites (Table 3). In order to identify the similarities and/or differences in the plant communities among the sites, CA ordination was carried out (Fig. 5). In CA ordination, axes 1 and 2 accounted for 12.1% (eigenvalue = 0.76) and 11.1% (eigenvalue = 0.70) of total variance, respectively. Relatively strong loadings on axis 1 were
Among the major dominant plant species in group I, we selected species which have over 10% coverage at a floor (Table 4).
The study sites represent three regions distributed by the mountain ranges which divide watersheds and cli-
[Table 3.] Dominant plant species recorded in abandoned paddy terraces.
Dominant plant species recorded in abandoned paddy terraces.
[Table 4.] Plant species over 10% coverage at a site in group I.

Plant species over 10% coverage at a site in group I.
remate zones. This indicates that each region might have different environmental condition. CA analysis confirmed that studied sites can be grouped into three, which were originally suggested by the relative location of the sites in South Korea. Such different climate could affect soil formation because a quantity of precipitation or a time and period of raining could be an important factor. Soil condition was a similar gradient among floors in the same abandoned paddy terraces but a quite difference among the locations. These environmental differences among the abandoned paddy terraces were confirmed by Duncan’s test. Among differences in environmental factors, K+, Ca2+, Na+, and Mg2+ contents were extremely high in site B. Site B was located near the east coast, so it could be affected by salt spray (Barbour and DeJong 1977) and shell crusts which were easily found in soil sample of this site. Water depth was also significantly different among the abandoned paddy terraces, not floors. In general, water level was low (less than 30 cm) and consistent through a year in all abandoned paddy terraces. However, site C showed a high physical gradient and water depth was quite different among the floors. Site C was disturbed by human activities and local government conserves this area as habitats of
A total of 141 species were investigated in sum of whole abandoned paddy terraces and 98 species were overlapped in all abandoned paddy terraces. 55 species were dominant plant species. CA analysis showed that all dominant species could be assigned to 4 groups. There are 10 common plant species having over 10% coverage of a floor in all abandoned paddy terraces. This indicates similar distribution pattern of dominant species despite site differences. We can expect 10 species in any between 10 and 15 years old abandoned paddy terraces in South Korea because tree layer was lower than 10% coverage and shrub layer was lower than 20% coverage (Lee 2006). Generally, abandoned paddy terraces in mountainous valley about 10 years old would show slow growth rate of vegetation because other abandoned paddy fields could be recovered with sprout placed in there but abandoned paddy terraces might be affected by newly entering seed (Bazzazz 1968, Harrison and Werner 1982, Inouye et al 1987, Lee 1995). The coverage of
This study confirmed that water depth was the most important factor for early successional stage in abandoned paddy terraces (Weiher and Keddy 1995). Also, diverse water depth induces diverse plant species because high environmental gradient could provide diverse conditions for diverse habitats (Ackerly 2003, Pausas et al. 2003). However, consistent deep water in a lentic wetland could limit the inhabitation of plant species (Miller et al. 2003).
There was the similar early development of plant community even though the abandoned paddy terraces showed different environmental conditions through they were divided into groups according to regions. In general, ecosystem development is affected by environmental conditions. In particular, climate and physico-chemical characteristics could have a strong effect on the development of vegetation structure (Mitsch and Gosselink 2007). However, development of plant community was similar in all abandoned paddy terraces in this study. This result might come from focusing dominant species which are determined by water level.
After abandoned, paddy fields in flat ground depending on irrigation systems might be dried. However, water regime in abandoned paddy terraces were mainly affected by around mountain stream and precipitation for water resource and levee and abandoned paddy terraces maintained nearly always wet condition and water level was maintained at a consistent level because of entire levees around arable fields (Park et al. 2006). Even though monsoonal climate experienced heavy rain during summer season, water depth in abandoned paddy terraces could keep at a constant level by slopes and levees due to flowing excessive quantity of water to downhill. Furthermore, inaccessibility to these locations could be also advantage to the disturbance by anthropogenic activities. For those reasons, abandoned paddy terraces could be showed lownutrient (conductivity of 30 to 40 μS/cm), which could relatively make interspecific competition weaken (Greulich et al. 2000). Very high nutrient levels were not to be favor of co-occurrence for species with contrasting nutrient requirements and nutrient level at abandoned paddy terraces were very low (Bornette et al. 1998). This might be accounted by diverse flora and the Red List species were recorded in abandoned paddy terraces (Uematsu et al. 2010). Thus, keeping consistent shallow water level and low nutrients condition were important characteristics to allow abandoned paddy terraces to develop similar level of plant community. It could be explained by the environmental sieve model (van der Valk 1981). Introduced species and survived species could be determined by environmental characteristics. A succession in wetlands began to open water and introduced species could be affected by characteristics of water regime. In particular, water depth was one of important factors to decide a process of early stage of succession (Mitsch and Gosselink 2007). Even though seeds of some species could be distributed over the wetlands by wind, animals or another means, they might be determined whether they could germinate and settle or not by environmental factors as filters that was assembly rules (Keddy 1992) and early settlement effect. After establishment, it seemed that what species would develop to the dominant plant community or not could be affected by another condition which could be competition between plant species or change topography or soil condition depending on plant community.
Early development of plant community in abandoned paddy terraces in South Korea could be determined by water regime rather than climate and physico-chemical characteristics of soil. This came from the similar water regime and location of abandoned paddy terraces, which play an important role in recruiting plant species. We conclude that early development of plant community in abandoned paddy terraces is similar in Korea even though the environmental conditions are different.