Cryptonemia J. Agardh (Halymeniaceae) is a foliose red algal genus comprising of 43 species that are distributed from warm to cold temperate regions of the world (Guiry and Guiry 2011). The genus Cryptonemia, typified with C. lomation (Bertoloni) J. Agardh, is distinguished by the well-developed stipe and / or midrib, highly refractive medullary cells, and bushy ampullar filaments with up to four orders (Chiang 1970, Womersley 1994). In Korea, two species of Cryptonemia are reported; C. lomation [as C. lactuca (C. Agardh) J. Agardh, Lee 2008; see Guiry and Guiry 2011 for synonymy] and C. tuniformis (= C. tunae-formis) (Bertoloni) Zanardini (Kang 1966). Both species are rarely found in Korea (Kang 1966, Lee and Kang 2001, Lee 2008).
Schizymenia J. Agardh (Schizymeniaceae) consists of 10 species that are distributed in temperate and tropical waters, ranging from the subantarctic islands to Alaska (Gabriel et al. 2011, Guiry and Guiry 2011). It is distin-guished by the presence of the gland cells resembling large, inverted tears, located terminally on cortical fila-ments (Dixon and Irvine 1977, Gabriel et al. 2011). Only Schizymenia dubyi (Chauvin ex Duby) J. Agardh is listed to date in the Korean inventory of marine algae (Kang 1966, Lee and Kang 2001, Lee 2008).
During floristic study of Korean marine algae, we found two unrecorded species; Cryptonemia rotunda (Oka-mura) Kawaguchi and Schizymenia apoda (J. Agardh) J.Agardh. In the present study, we analyzed rbcL sequences of both species together with previously recorded S. dubyi through extensive collections along the coasts of Korea, observing details of morphology. Published sequences of species of Cryptonemia and Schizymenia (Kawaguchi et al. 2004, Gabriel et al. 2011, Huisman et al. 2011) allow us to directly compare our results with others from these two genera as a whole and with putative relatives. We found that both Cryptonemia and Schizymenia have the deep taxonomic problem and the lack of update in many areas of the world.
Field collections of Cryptonemia and Schizymenia were made at many locations in Korea (Table 1). Materi-als were collected from intertidal zone or subtidal zone
using SCUBA diving. Tissues were sectioned with a freez-ing microtome (FX-80; Yamato Kohki Industrial Co. Ltd., Saitama, Japan). Photographs were taken with a DP-71 camera (Olympus, Tokyo, Japan) attached to a micro-scope (BX 51; Olympus). Voucher specimens are housed at the herbarium of Chungnam National University, Dae-jeon, Korea (CNUK).
Forty-one specimens were available for extraction of DNA: 10 for C. rotunda, 7 for S. apoda, 24 for S. dubyi (Ta-ble 1). DNA extraction, polymerase chain reaction ampli-fication, and sequencing follow Boo et al. (2010). Primer pairs for amplification and sequencing of rbcL were as follows: rbcLF145-rbcLR898 and rbcLF762-rbcLR1442 (Kim et al. 2010).
Maximum likelihood (ML) phylogenetic analysis of rbcL was performed using the GTR + Γ model imple-mented in RAxML software (Stamatakis 2006). We used 200 independent tree inferences with the ‘number of run’ option, with default optimized subtree pruning and re-grafting rearrangement and 25 distinct rate categories to identify the best tree. Statistical support for each branch was obtained from 1,000 bootstrap replications using the same substitution model and RAxML program settings.
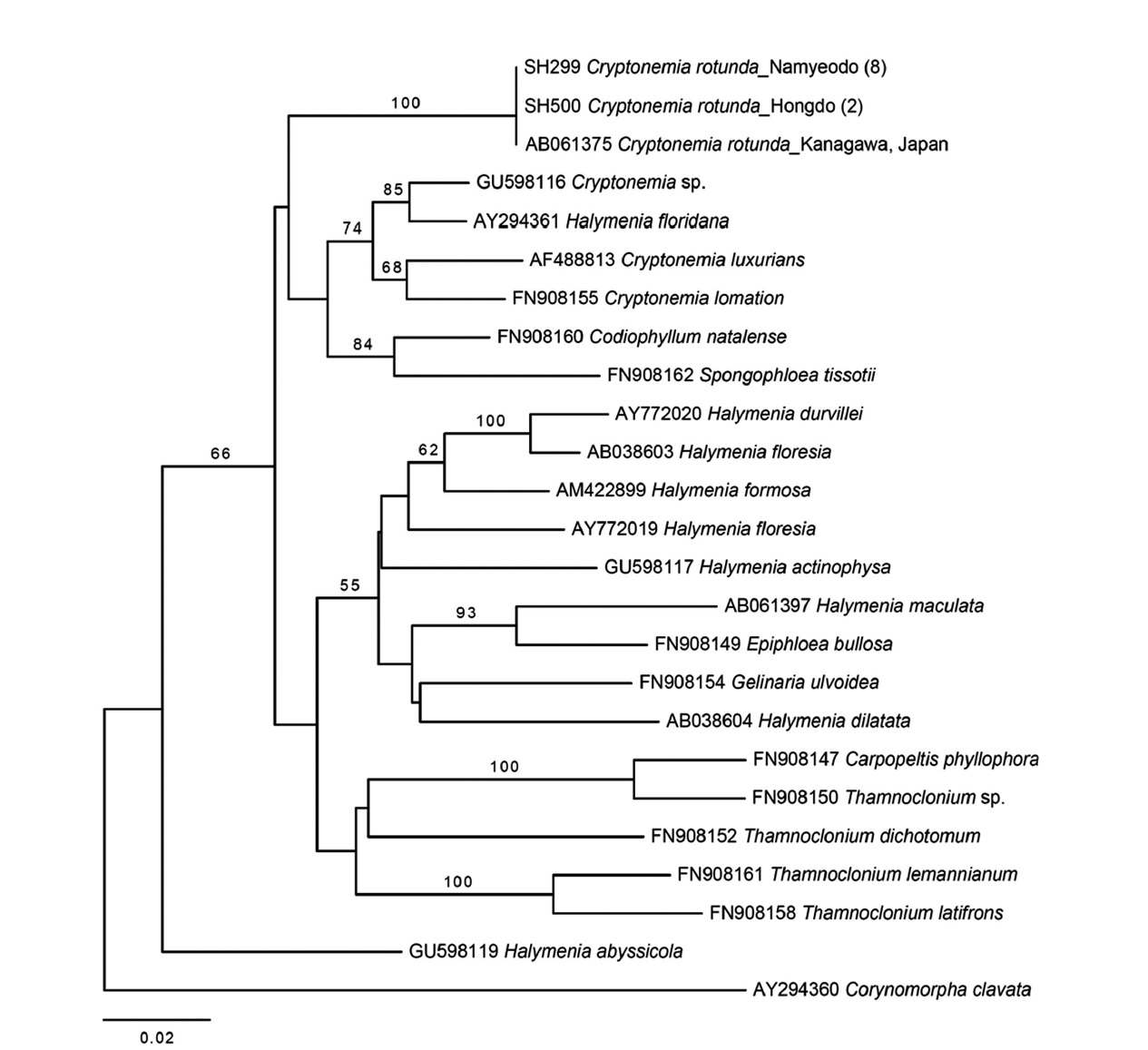
Molecular analyses. Thirty-three rbcL sequences con-sisting of 23 taxa of Halymeniaceae including ten new sequences of C. rotunda analyzed in the present study were aligned with 1,129 bp. All Cryptonemia specimens collected from Hongdo and Namyeodo belong to a single species consistent with the GenBank sequence of C. ro-tunda from Japan (Fig. 1).
Plastid rbcL tree revealed that the genus Cryptone-mia was not monophyletic (Fig. 1). C. rotunda was not monophyletic with the other members of the genus. The pairwise divergence between C. rotunda and Halymenia floridana was 5.2%, while C. rotunda was in distance of 5.6-6.3% from other members of the genus.
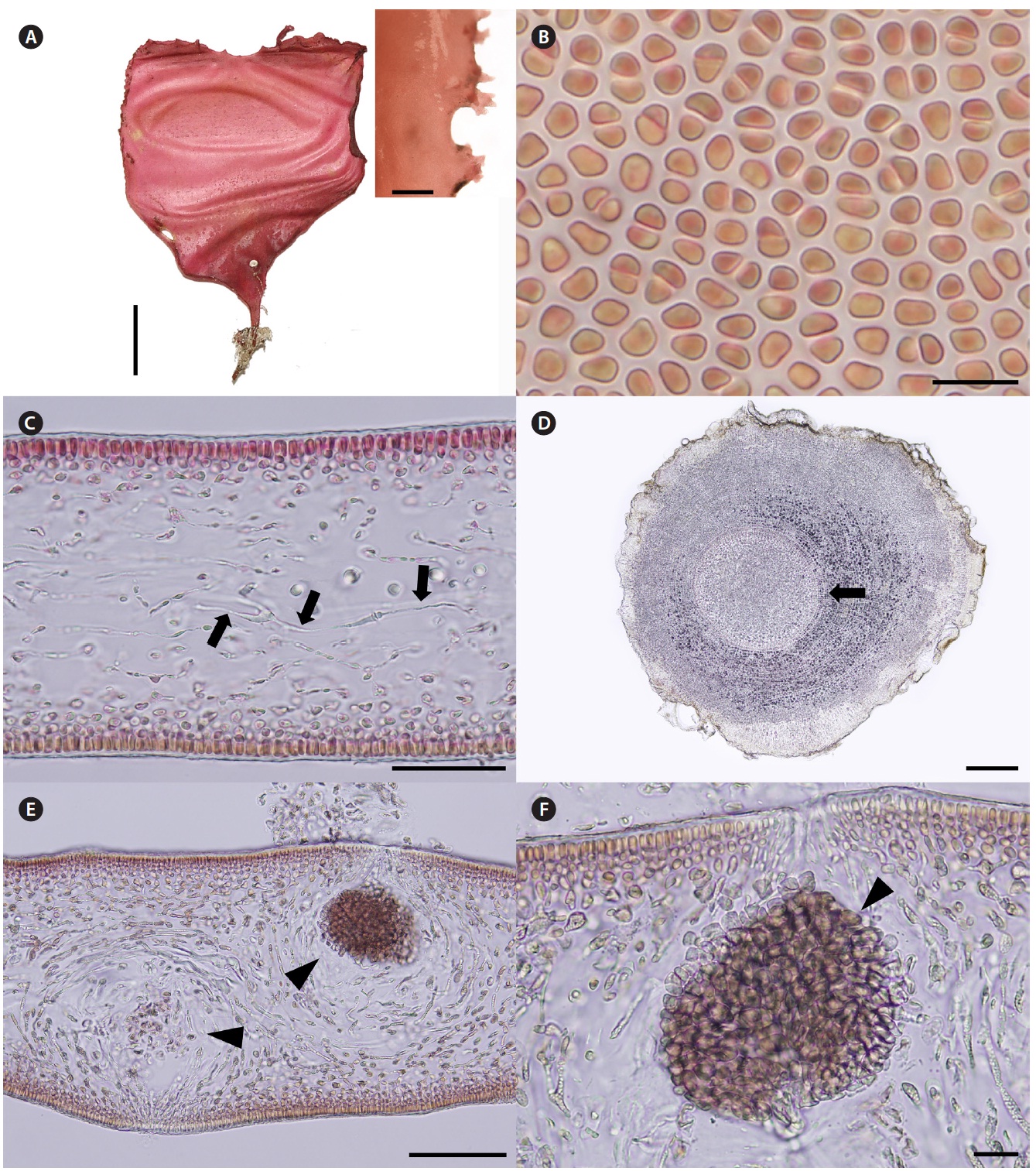
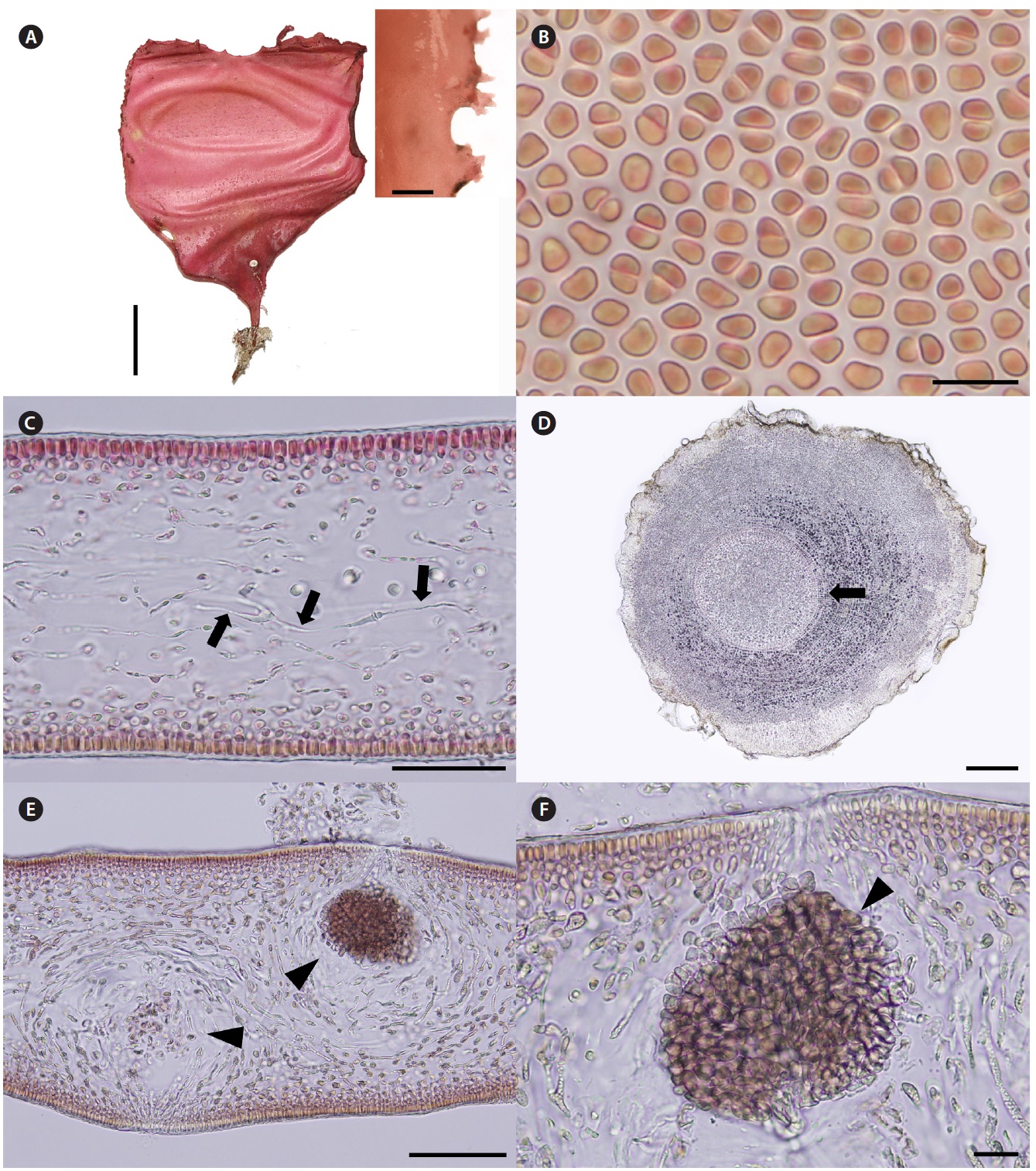
Korean name. 둥근바다선인장 (신칭) (Fig. 2A-F)
Description. Thalli are medium to dark red, membra-nous, circular or ellipsoid, rarely irregularly shaped, up to 8 cm in length and 4 cm in width, expanding from a stipe (Fig. 2A). Blades are thickened along the margin and beset with numerous, small protuberances, looking toothed (Fig. 2A), up to 200 μm thick (margins 300-370 μm thick). Cells at the surface of blade are angular (Fig. 2B). Tissue consists of cortex and medulla (Fig. 2C). Corti-ces are compact with 2-5 layers of cortical cells and 20-30 μm thick. Medulla are 120-180 μm thick, of entwined re-fractive filaments 1-4 μm in diameter (Fig. 2C). Holdfasts are discoid, 1-3 mm across, bearing stiff stipe which have several growth rings in cross section (Fig. 2D).
Female reproductive structures are scattered around upper side of the blade. Cystocarps (Fig. 2E & F) are sub-globose, 80-100 μm across, and swollen on the surface of blades with small ostiole. Both male plants and tetraspo-rangial plants were not found.
Thalli grow on rocks at a depth of 14-18 m in the sub-tidal zones. Thalli are larger in July than in November.
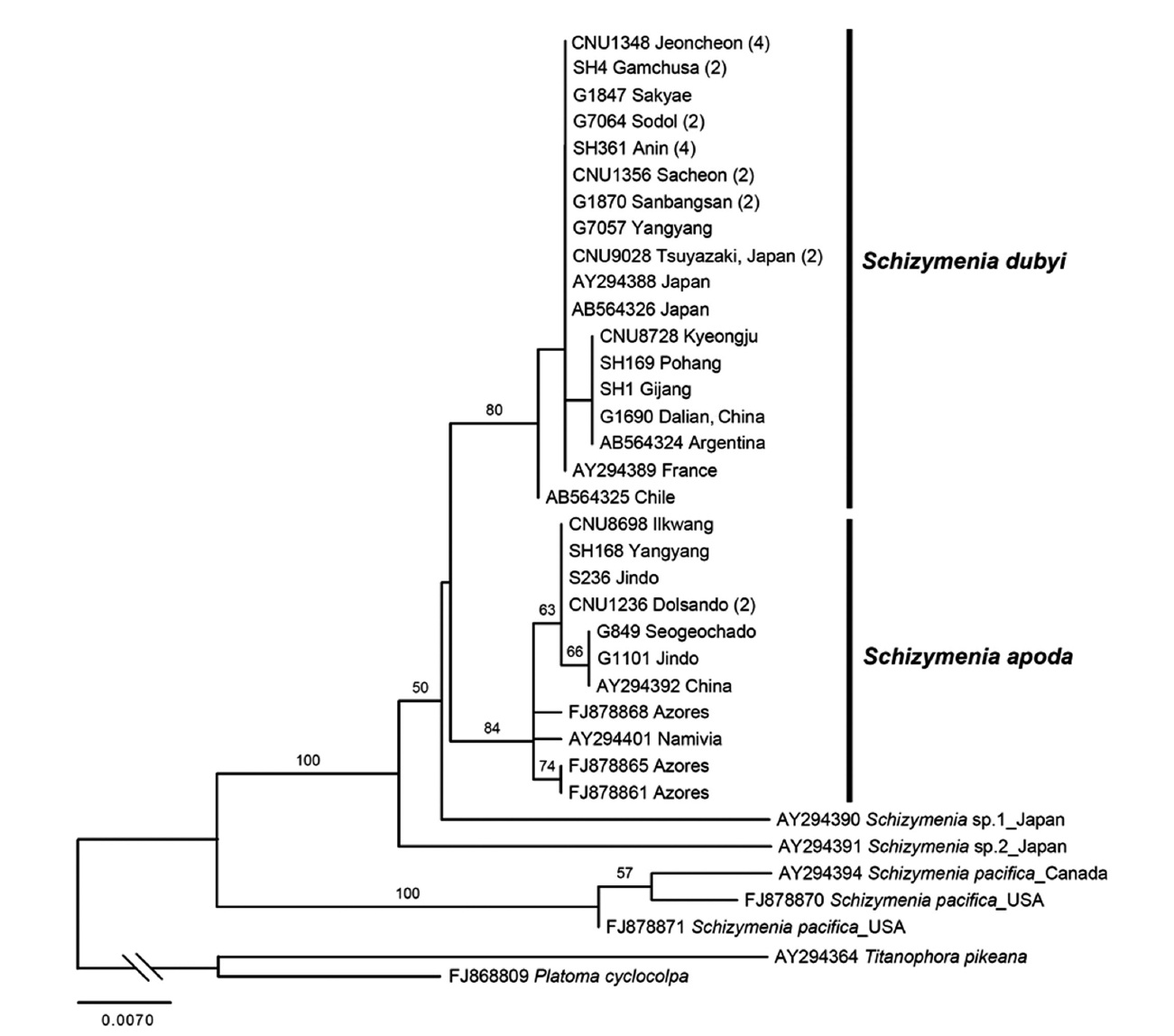
Molecular analyses. A total of 48 rbcL sequences of Schizymeniaceae including 31 new sequences were aligned with 503 bp. Seven specimens of Korean S. apoda clustered with those from China, the Azores, and Na-mibia (Fig. 3). Twenty-four specimens of S. dubyi from Korea clustered with those from China, Japan, Argentina,
France and Chile.
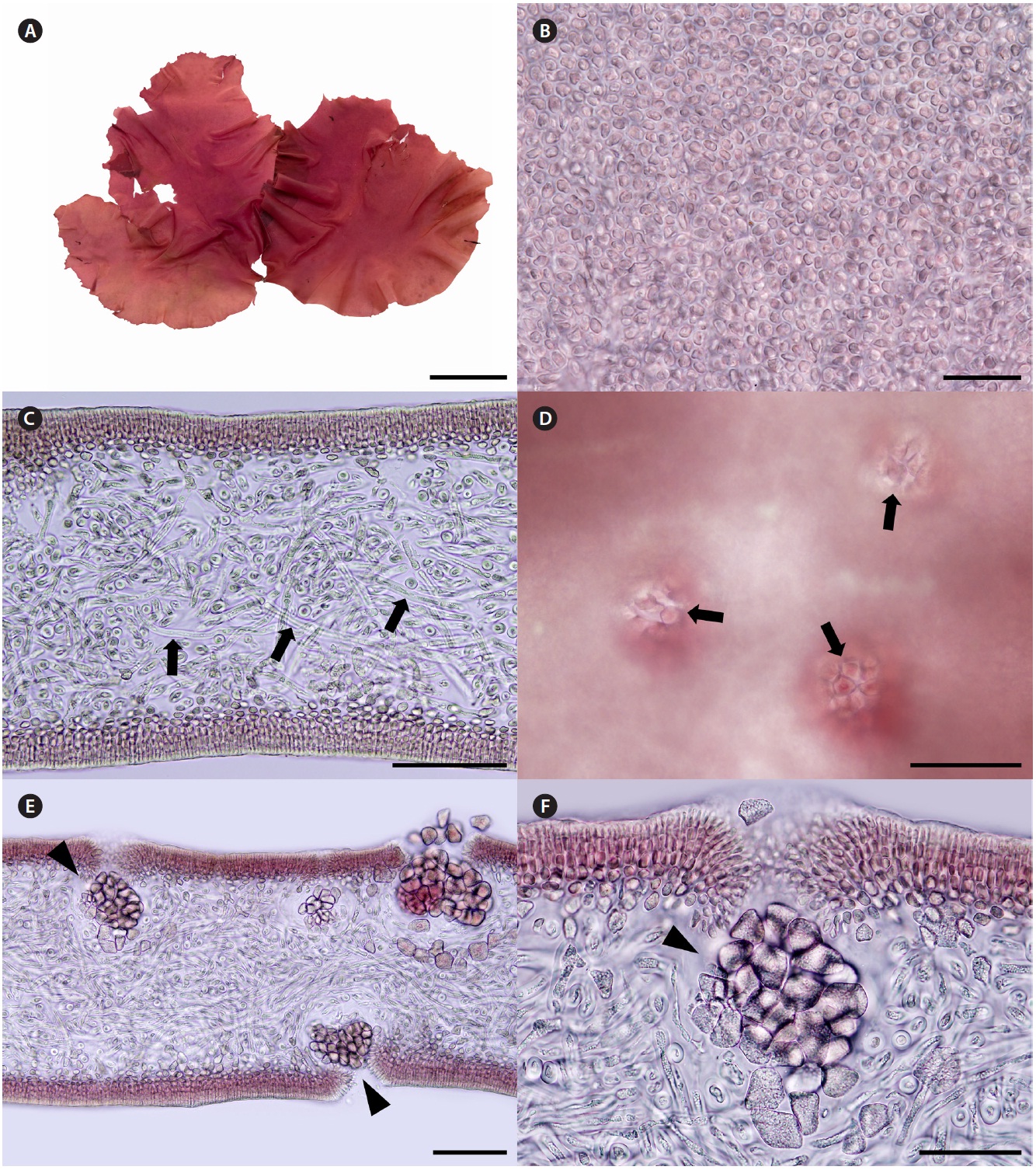
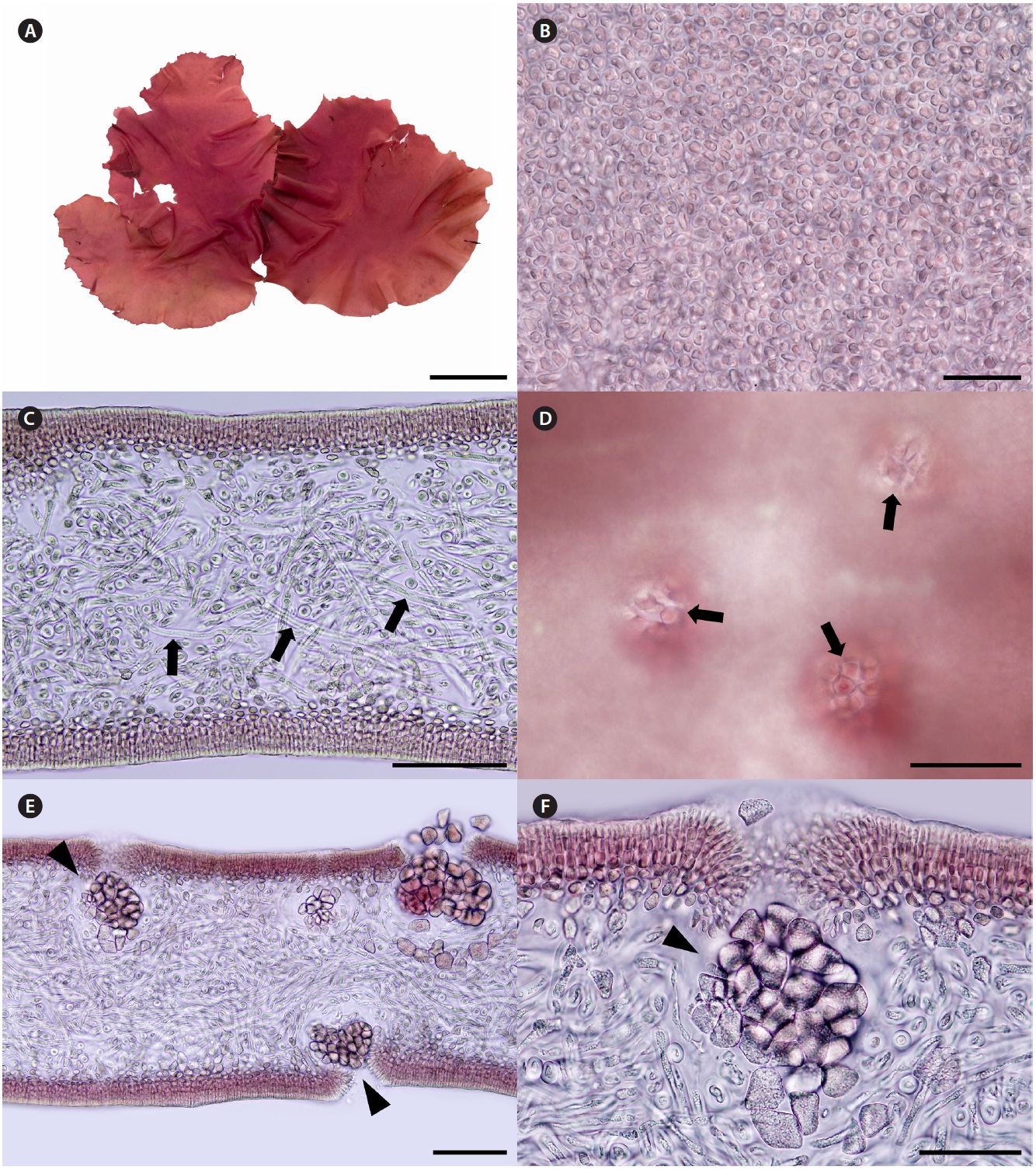
Korean name. 자루갈래잎(신칭) (Fig. 4A-F)
Description. Thalli are bright to dark red, soft fleshy, circular or ellipsoid with undulated margins, up to 32 cm in length and 15 cm in width (Fig. 4A). Cells at the sur-face of blade are irregularly elongate (Fig. 4B). Tissues are compactly composed, and 300-330 μm thick (Fig. 4C). Cortices are 5-8 layers of cortical cells and 40-60 μm thick. Medulla are composed of thin filaments that are densely interwoven with one another, and 200-240 μm thick.
Female reproductive structures are scattered over en-tire blade, and easily distinguished by visible ostioles (Fig. 4D). Cystocarps (Fig. 4E & F) are subglobose, 40-80 μm across. Spermatangia and tetrasporangia are not found.
Thalli grow on the rock with coralline algae at intertidal zones. They were collected in February, March, and April from 2008 to 2011.
Plastid rbcL sequences of ten specimens of C. rotunda collected from the subtidal in Hongdo and Namyeodo is-lets on the southeastern coast of Korea were identical with that (AB061375) of a Japanese specimen. Thalli have sub-stantial stipe and circular, membranous blade, and highly refractive cells in medulla. Morphology and anatomy of the Korean specimens correspond well to the description of Okamura (1936, as Halymenia rotunda Okamura) and Kawaguchi (1993). This is the third species in the Korean Cryptonemia inventory, together with C. lomation and C. tuniformis. C. lomation and C. tuniformis from Korea are different from C. rotunda in having midribs (Kang 1966, Lee 2008). C. lomation occurs only in Udo, Jeju Island (Lee 2008), while C. tuniformis has not been reported after Kang (1966, 1968) who described it based on a single frag-ment cast up ashore in Jijindo, now included to Yeochon city, on south coast. Molecular analysis of C. lomation and C. tuniformis is necessary to confirm their genetic identities because both species are mainly distributed in Europe (see Guiry and Guiry 2011). The topology of our rbcL tree is similar to that of previous studies on the fam-ily Halymeniaceae (Kawaguchi et al. 2004, Huisman et al. 2011). However, our tree reveals that Cryptonemia is not monophyletic and C. rotunda is distantly related to other species of Cryptonemia. More sampling is necessary for the urgent revision of the genus Cryptonemia.
The second species added to the algal inventory of Ko-rea is S. apoda, which has, to date, been recorded in South Africa, Europe, and China (Gabriel et al. 2011). Thalli of Korean S. apoda are similar to those from the Azores (Gabriel et al. 2011). Thalli are foliose, simple or broadly lanceolate blades with irregular lobes and proliferations from the margins, sometimes undulate, soft fleshy, and slippery, up to 32 cm tall. Cystocarps on the blade surface have ostioles. In the present study, we found S. apoda in the same (e.g., Namaehang in Yangyang) or nearby lo-cations where S. dubyi was recorded in Korea (see Table 1). S. dubyi is very common from Sokcho to Guryongpo on the east coast, and Busan to Heuksando on the south coast (Kang 1966), and Jeju Island (Kang 1966, Lee 2008). The mixed occurrences of S. apoda with S. dubyi in Korea imply that S. apoda thalli have likely been misidentified as morphological variants of S. dubyi. Our results support the suggestion of Gabriel et al. (2011) that at least several of the Japanese S. dubyi records (e.g., Yoshida 1998) are likely to be S. apoda.
All specimens of Korean S. apoda always formed a clade with those from the Azores, China, and Namivia in rbcL phylogeny (Fig. 3). However, genetic diversity with-in S. apoda is likely higher than S. dubyi (up to 0.6% for S. apoda, 0.4% for S. dubyi). All specimens of S. dubyi in Korea formed a clade with those from Argentina, Chile, China, France and Japan. S. apoda and S. dubyi formed a sister clade.
Together with the previous study by Gabriel et al. (2011), our result demonstrates that both S. apoda and S. dubyi have disjunct global distributions: China, Korea, the Azores, Namivia for S. apoda and Argentine, Chile, China, France, Japan, and Korea for S. dubyi. It is interest-ing to study whether their disjunct distributions can be explained by either natural dispersal or anthropogenic transport, using more taxon sampling and more variable molecular markers such as mitochondrial cox1.