Recently, Ueda and Nagai (2009) described three new spe-cies from Ariake Bay of Kyushu, Japan. In the remarks sec-tion of one of their three new species,
As ‘
Herein, I redescribe
Material examined in the present study was obtained from estuaries and salt marshes in South Korea since August, 1984, and stocked in the Department of Biological Science, Daegu University. Samplings were made using a dipnet or a coni-cal plankton net with a 64 ㎛ mesh.
Specimens were dissected and mounted in lactophenol on a Cobb slide, after treatment in a solution of 5% glycerin-95% ethyl alcohol for 1-2 days. Dissection was performed using two needles made from 0.3 mm diameter tungsten wire by electrolysis. Mounted specimens were observed using a differential interference contrast microscope (Olympus BX-51, Japan) equipped with Nomarski optics. Measurements were done using a digital camera equipped to mount on the microscope (Cool SNAP 5.0M; Roper Scientific, USA) and QCapture Pro ver. 5.0 calibration software (Media Cybernet-ics, USA).
Voucher specimens have been deposited in the National Institute of Biological Resources, Incheon, Korea, and in the specimen room of the Department of Biological Science, Daegu University, Korea.
Morphological terminology follows Huys and Boxshall (1991). The Sewell’s style (1949) is adopted for the armature formula of the swimming legs, in which spines are denoted by Roman numerals and setae by Arabic numerals. Abbre-viations used in the text and figure legends: A1, antennule; A2, antenna; Fu, caudal ramus; L/W, length to width ratio; P1-P5, legs (pereiopods) 1-5; enp 1-3 or exp 1-3, the first to third endopodal or exopodal segments of each leg.
Order Cyclopoida Burmeister, 1835
Family Cyclopidae Dana, 1846
Subfamily Halicyclopinae Kiefer, 1927
Genus Halicyclops Norman, 1903
1*Halicyclops continentalis Ueda and Nagai, 2009 (Figs. 1-3)
1*
Material examined. Korea: 3♀,♀ Chungcheongnam-do: Dangjin-gun, Songsan Reservoir, 36? 8′33′′N, 126? 3′20′′E, 5 May 2010, Chang CY, Lee J; 6♀♀, Dangjin-gun, Songak-eup, Bogun-ri, former salt farm, 36? 5′22′′N, 126? 6′50E′′, 1.5-2.1 psu, Sep, Oct 2010, Moon SD; 4♀,♀ Seosan-gun, Palbong waterway, 36?8′48′′N, 126?20′08E′′, 23 Jul 2009,Chang CY, Lee J; 2♀,♀ Seocheon-gun, detention reservoir at Busa Tidal Embankment, 36? 11′39′′N, 126? 32′24E′′, 15 Jul 2003, Chang CY, Lee J.
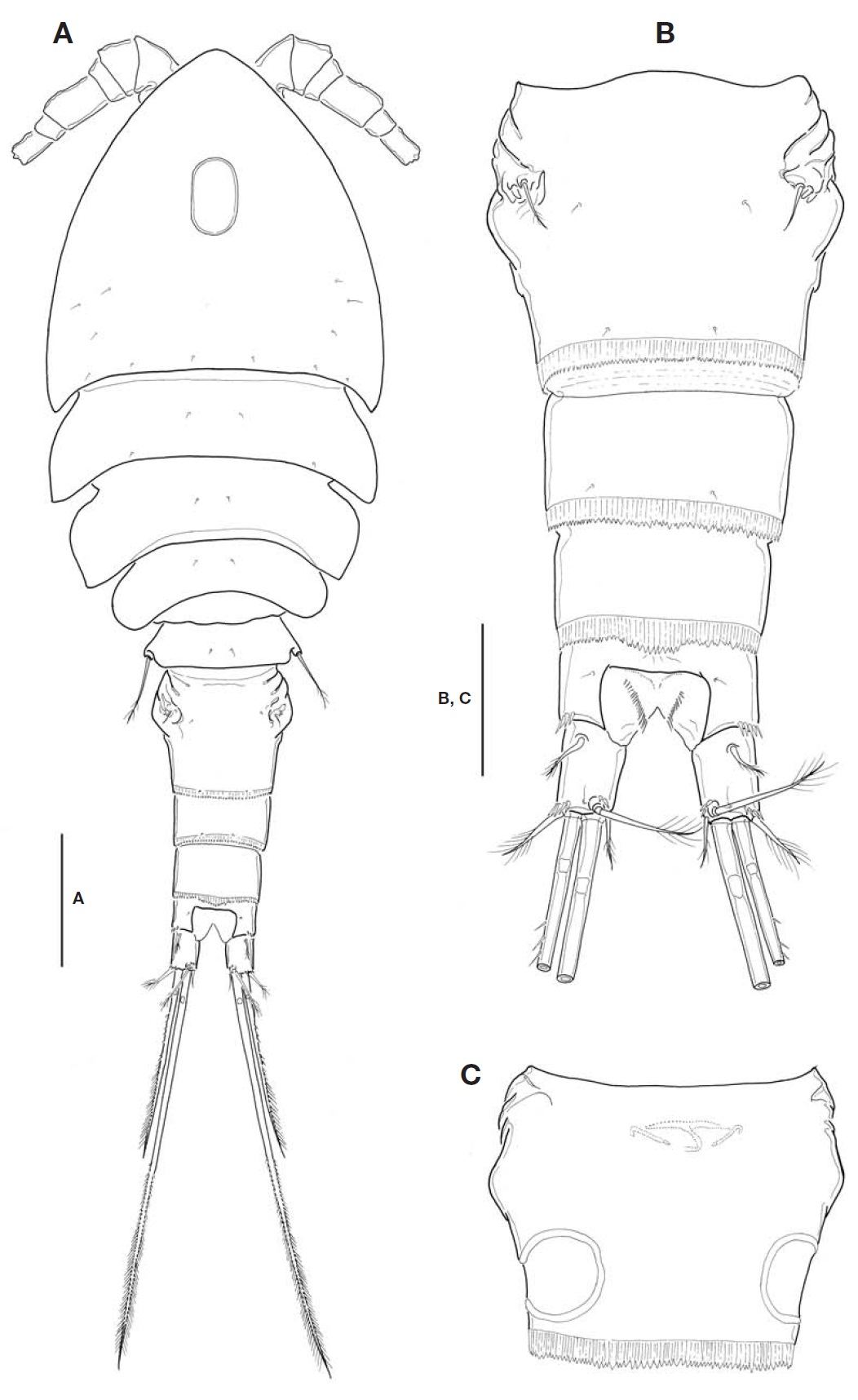
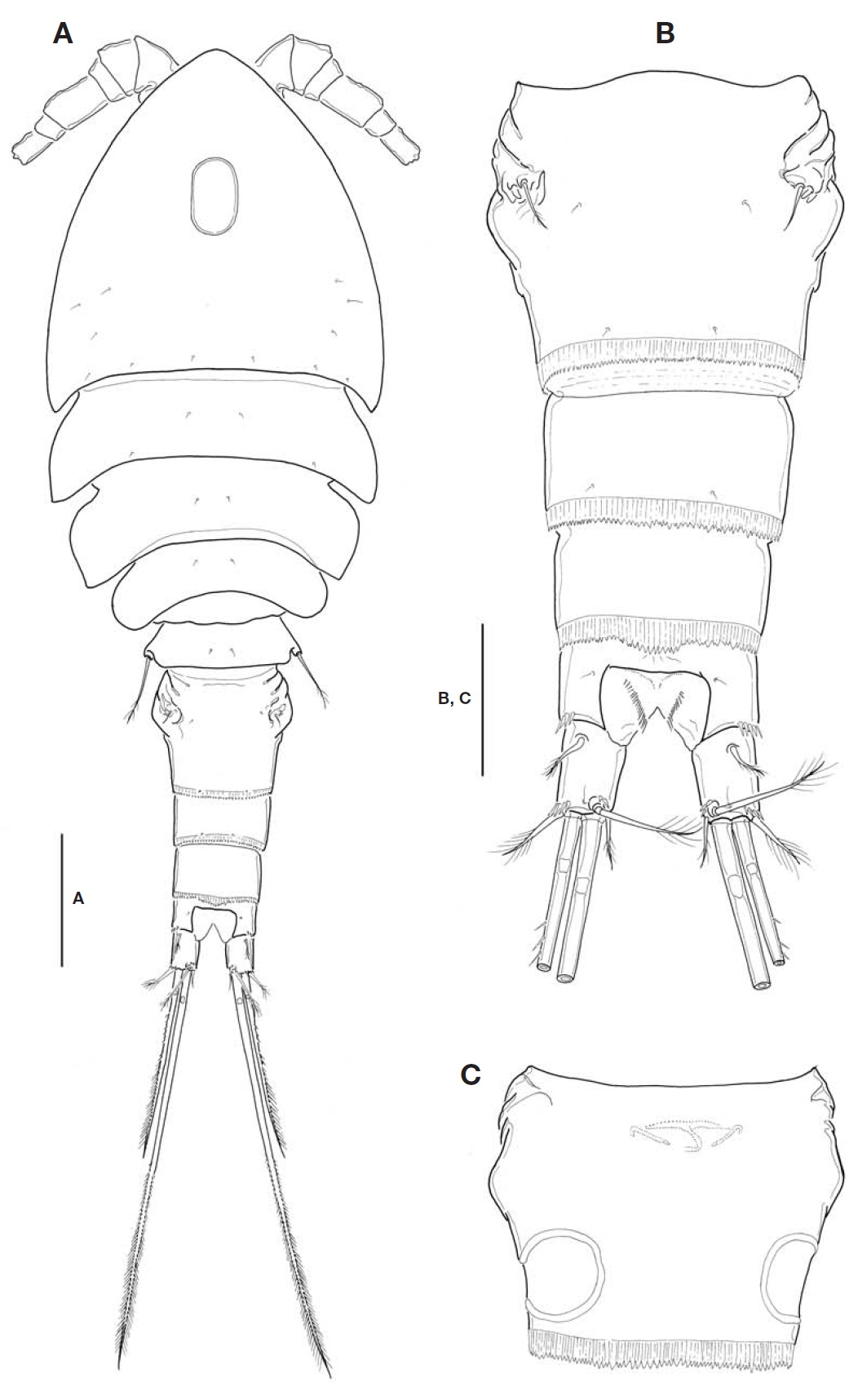
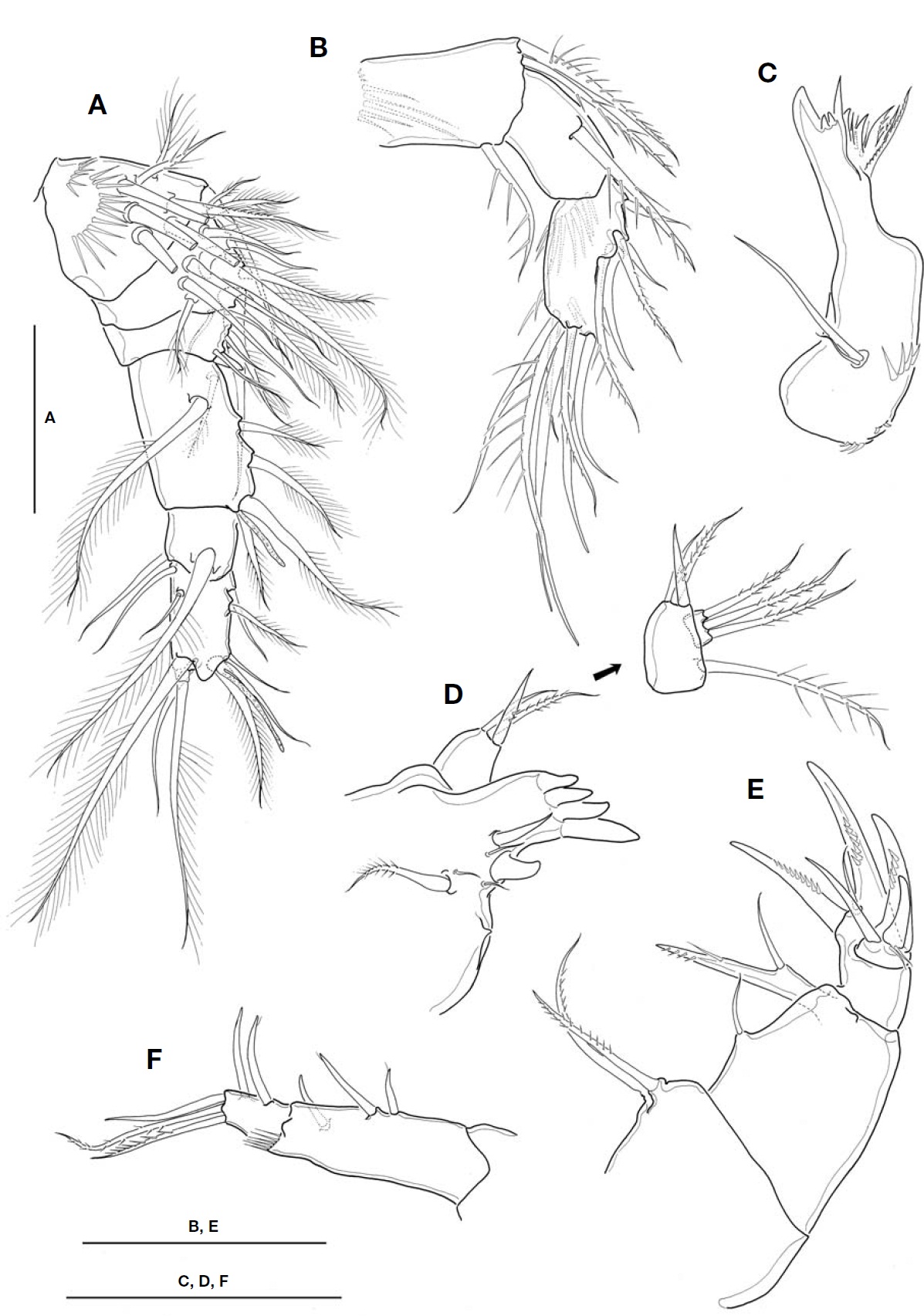
Description. Female: Body (Fig.1 A) relatively small, mean 683.6 ㎛ long (range, 664. 2 to 698.4 ㎛, n=12), excluding caudal setae; maximum width at posterior end of cephalo-thorax, 252.8 ㎛; L/W about 2.7. Body slightly flattened dorso-ventrally. Prosome comprising cephalothorax incorporating first pedigerous somite and 3 free pedigerous somites; poste-rior margins of prosomites nearly smooth, not serrated; outer distal corners of first three prosomites weakly expanded pos-teriorly. Cephalothorax strongly protruded anteriorly, so somewhat triangular in general appearance; L/W about 1.1; ellipsoidal integumental window (or median depression) pre-sent mid-dorsally; 12-18 sensilla scattered on dorsal and lat-eral surfaces. Rostral expansion rarely visible from dorsal view. Fifth pedigerous somite incorporating basis and endo-pod of P5, bearing a basal seta outer posterior corner of dor-sal surface. Genital double-somite (Fig.1 A-C) much wider than long (L/W 0.63, at level of lateral expansion); both sides of anterior part expanded laterally, lacking spinous processes; lateral expansion somewhat rectangular in general with its corner not pointed but obtuse or round; in dorsal view, strong-ly sclerotized wrinkles just ahead lateral expansion armed with 3 appendages representing P6 (Fig.1 B); genital area simple, with a big median copulatory pore anteriorly; with paired cuticular recesses ventrolaterally at posterior half of genital double-somite (Fig.1 C). Mid-dorsal hyaline fringe of preanal somite protruding posteriorly, with 12-15 teeth along posterior margin, slightly not extending over anal oper-culum (Fig.1 B).
Fu (Fig.1 B) not elongate, about 1.38 times longer than wide; divergent posteriorly; armed with spinules at outer dis-tal margin of ramus and near base of outer caudal seta. Lat-eral caudal seta lying proximally and dorsally. Inner caudal seta 0.7 times as long as ramus. Outer caudal seta slightly shorter than ramus. Dorsal caudal seta arising from shallow protuberance, 1.5 times longer than ramus. Small cuticular tube with a minute pore present outside the protuberance. Both inner and outer terminal caudal setae with breaking planes; outer terminal caudal seta ornamented heterogeneous-ly on its outer margin: proximal quarter bare, second quarter pinnate, distal half plumose.
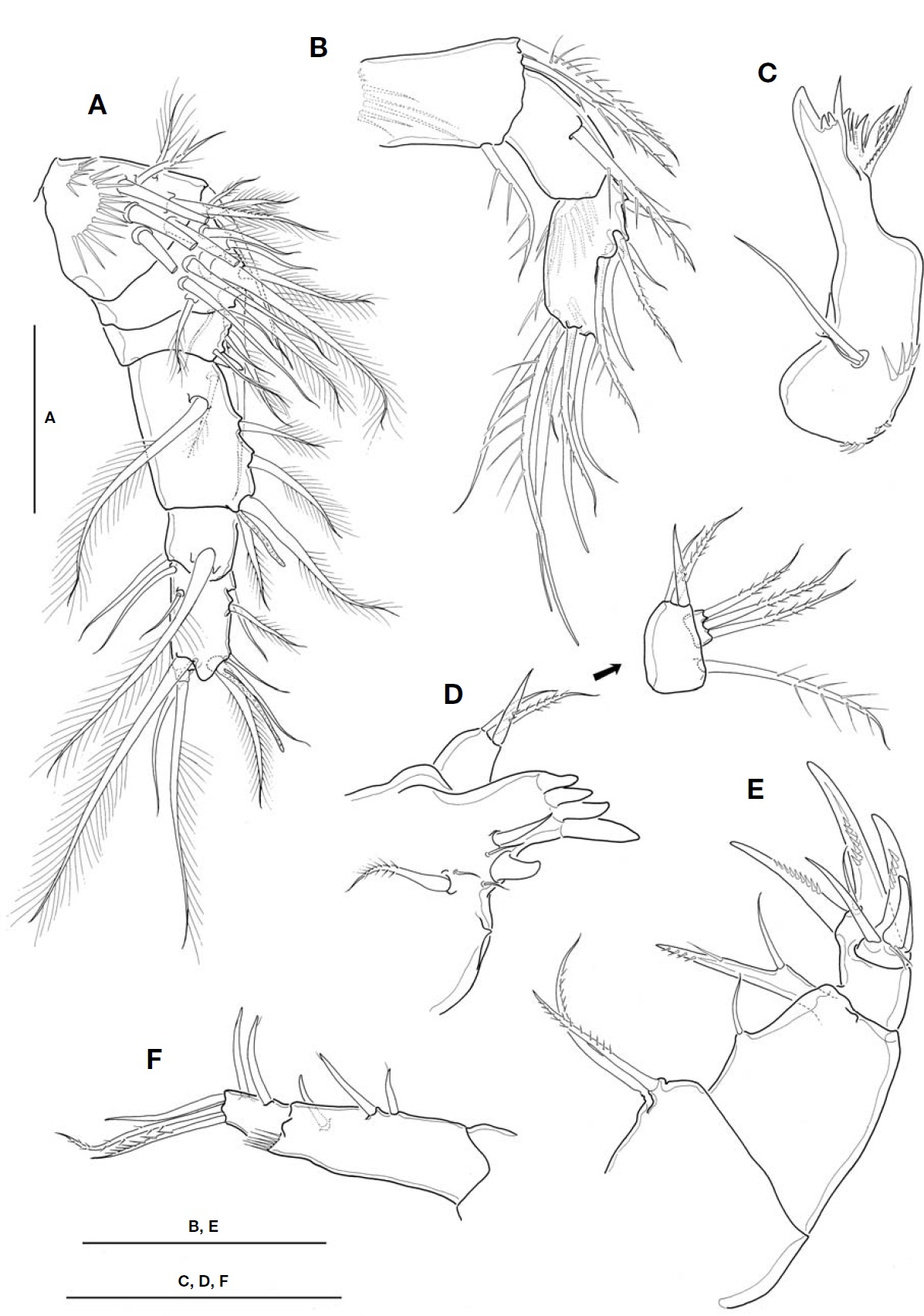
A1 (Fig.2 A) not reaching to halfway of cephalothorax; 6-segmented. Fourth segment 1.4-1.5 times as long as wide; last segment about 1.9 times longer than wide. Aesthetasc each arising from fourth and last segment distally. Seta for-mula: 8, 12, 5, 6 (+1 aesthetasc), 2, 10 (+1 aesthetasc).
A2 (Fig.2 B) 3-segmented, comprising coxobasis and 2-segmented endopod. Coxobasis ornamented with 1 setule row proximally, armed with 2 pinnate inner distal setae, and 1 outer distal seta representing exopod. First endopodal seg-
ment bearing 1 pinnate inner seta, with smooth margins lack-ing spinules. Second endopodal segment 1.8 times as long as wide, about 1.1 times longer than first endopodal segment; ornamented with 1 spinule row on caudal face proximally; armed with 5 lateral and 7 apical setae.
Mandible (Fig.2 C), with well-developed coxal gnathobase, armed with 6 strong teeth, 4 slender spiniform setae and 1 stout spine along cutting edge, flanking 1 outer distal pinnate seta. Palp very reduced, represented by 2 naked setae on small protuberance, of which shorter one small, about 1/3 times as long as longer one.
Maxillule (Fig.2 D), praecoxal arthrite bearing 4 strong tooth-like spines distally; 7 elements with various shapes present along inner margin, including 1 small proximalmost projection. Palp 2-segmented; coxobasis bearing 1 pinnate and 1 spiniform and 1 naked setae inner distally, plus 1 outer pinnate seta represent-ing exopod; distal segment, representing endopod, with 3 long plumose setae.
Maxilla (Fig.2 E) 4-segmented, comprising praecoxa, coxa,basis and 1-segmented endopod. Praecoxal endite with 2 pin-nate setae. Coxa with 1 naked seta representing proximal endite; distal endite movable, forming 1 bisetose lobe com-pletely fused with naked seta proximally. Basis forming 2 strong claw-like spines with 1 naked seta between them basal-ly. Endo-pod carrying 5 elements of 2 claw-like spines, 1 naked spiniform seta and 2 minute slender setae.
Maxilliped (Fig.2F) 2-segmented, comprising protopod and completely defined endopod; protopod about 3 times longer than endopod. Protopod with 2 naked spiniform setae proximally and 1 spiniform seta distally, representing endites; 1 setule row present on outer distal corner of protopod. Endo-pod bearing 5 setae in total, comprising 2 inner setae, 1 api-cal, 2 subapical pinnate setae.
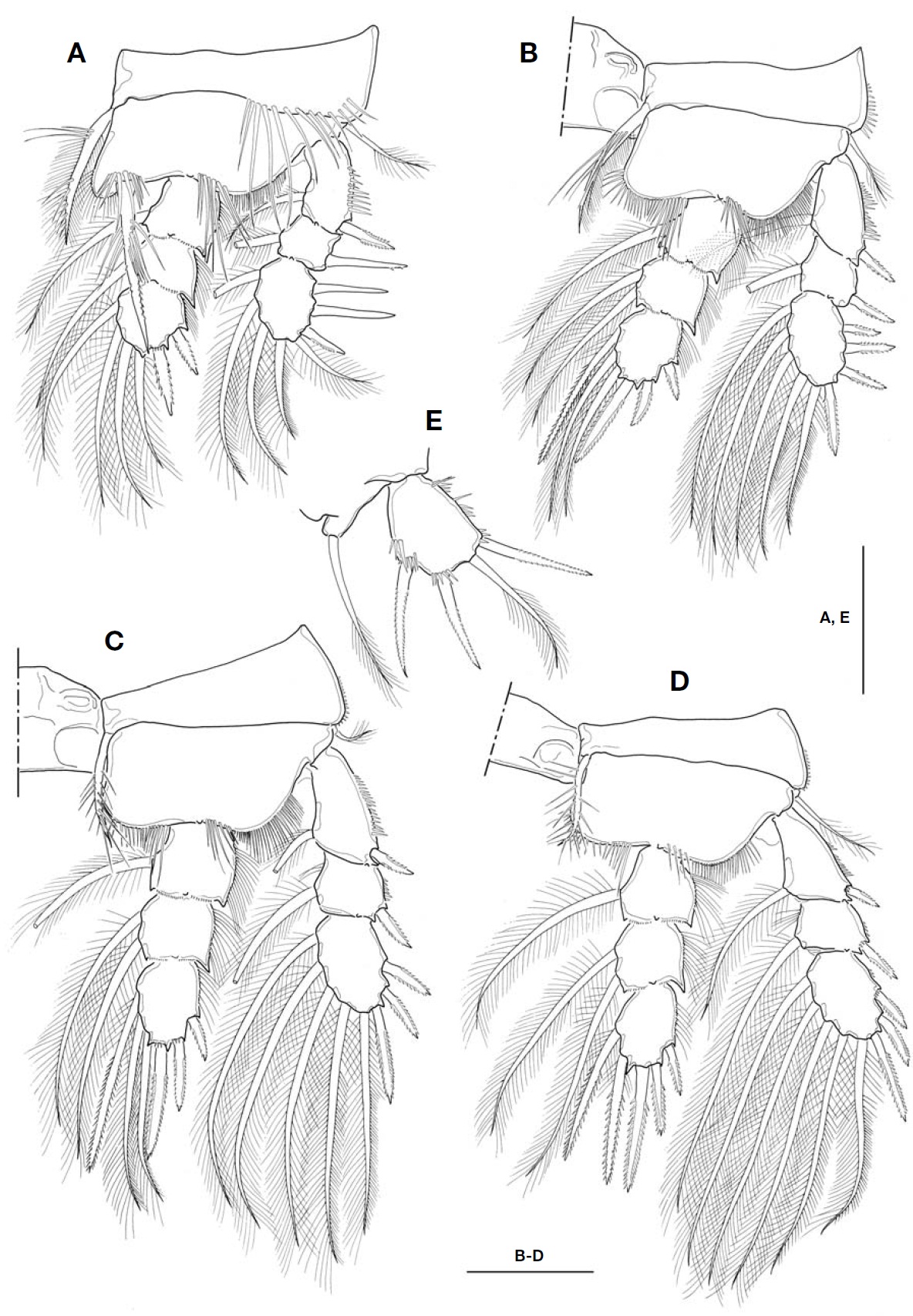
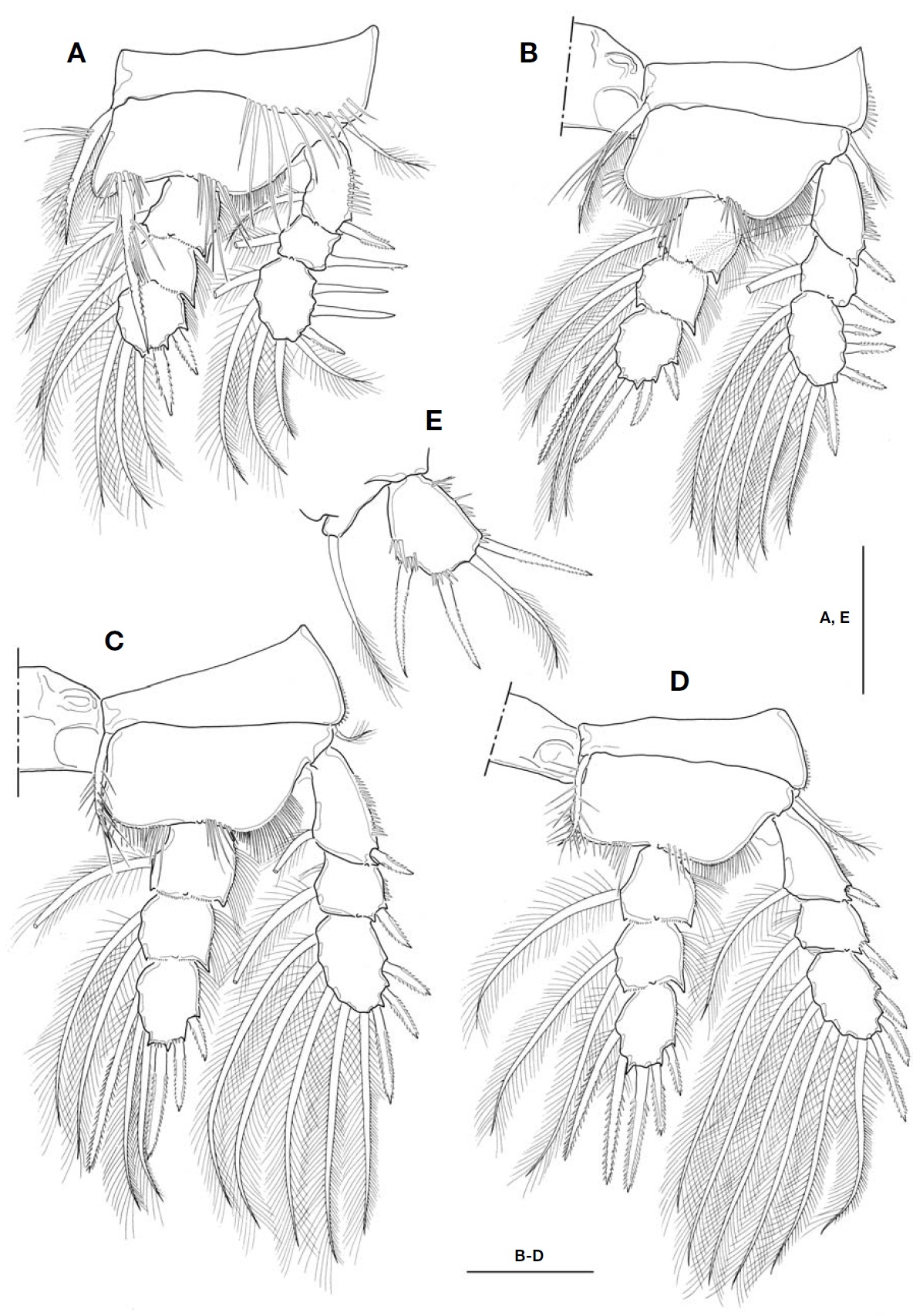
P1-P4 (Fig.3 A-D) biramous, both rami 3-segmented. Coxal setae stout, pinnate. Spine formula of 3, 4, 3, 3. Seta/spine arma-ture of P1-P4 as follows:
P1 coxa 0-1 basis 1-1 exp I-1; I-1; III, 1, 4 enp 0-1; 0-1; II, 1, 3
P2 coxa 0-1 basis 1-0 exp I-1; I-1; IV, 1, 4 enp 0-1; 0-2; II, I, 3
P3 coxa 0-1 basis 1-0 exp I-1; I-1; III, 1, 4 enp 0-1; 0-2; II, I, 3
P4 coxa 0-1 basis 1-0 exp I-1; I-1; III, 1, 4 enp 0-1; 0-2; II, I, 2
P1 (Fig.3 A), intercoxal sclerite with hairs along distal margin of both lateral lobes; inner distal seta on P1 basis stout, ornamented with long setules proximally and secon-dary spinules distally, and very long, slightly beyond pos-terior end of enp 3; distal two spines on exp 3 strikingly longer, about 1.8-1.9 times longer than proximal one. P2-P4, intercoxal sclerite bare along distal margin of both lateral lobes; enp 2 with 2 inner setae. P4 enp 3 (Fig.3 D) about 1.7 times longer than wide; both inner setae apparently spiniform, but proximal quarter of each seta retained plumose; apical spine 1.6 times longer than enp 3, and about 1.8 times longer than outer apical spine.
P5 (Fig.3 E), basis and endopod completely incorporated to fifth pedigerous somite; basal seta inserted on small pro-tuberance arising from dorsolateral corner of somite. Exopod not elongate, about 1.2-1.3 times longer than wide; inner margin nearly straight, ornamented with spinule rows; inner apical spine longest, only 1.2 times as long as exopod, and 1.1 times longer than outermost spine; apical plumose seta 1.3-1.4 times longer than exopod. P6 (Fig.1 B) indistinct,represented by small genital operculum armed with 3 ele-ments of 2 minute spinous projections and 1 long seta.
Male: Unknown.
Ecology. Specimens were collected from marshes with low salinity at the innermost part of reclaimed region, and from a few estuaries discharging into the west coast of Korea.
This species co-occurred with
Distribution. Japan (Ariake Bay, Kyushu), China (the Yel-low Sea coast between Tianjin and Guangzhou), and Korea (the west coast).
Remarks. At present, according to the number of spines on the distal exopodal segments of P1-P4 (spine formula), seven morphological groups of species are pointed out in the genus
The original description did not place weight on the dorsal integumental window (or median depression) on cephalotho-rax, which has been known in only three species in the genus
Korean specimens show minor discrepancies with the ori-ginal description: (1) lateral expansion is angular in general appearance, but not acute or rather round at its tip (vs. form-ing nearly angular); (2) Fu is a little more elongate than in the original description (L/W, 1.38 vs. 1.1-1.2); (3) dorsal caudal seta is 1.53 times longer than Fu (vs. 2.0), and inner caudal seta 0.7 times as long as Fu (vs. about same); (4) the fourth segment of A1 is relatively long (L/W, 1.4-1.5 vs. 1.3); and (5) P5 exopod is a bit elongate (L/W, 1.25 vs. 1.1). The paired cuticular recesses at the ventrolateral part of the pos-terior half of genital double-somite (cf. Fig.1 C) were not mentioned in the original description and must have been omitted in their Fig.2 C, which was emphasized as one of the decisive characters in the subgenus system by Karanovic (2006, 2008).
Ueda and Nagai (2009) regarded the
In the Ariake Bay, the type locality of this species, three brackish calanoid species (
Korean name: 1*대륙짠물검물벼룩 (신칭)