In Korea, base metal mines producing Cu-Pb-Zn ores are distributed almost over all of the country and were actively operated until the early 1980s. However, since then, base metal production has declined and most mines closed mainly due to economic reasons (Lee and Lee 2001). Upon closing of the mines, improperly disposed of mineral waste piles and untreated mine drainage have become important sources of heavy metals in the environment. In particular, lead (Pb) contamination increased substantially with industrialization of the forest(Shotyk et al. 1998). Forest ecosystems are particularly sensitive to Pb pollution from atmospheric deposition, as their canopies have a high capacity to intercept aerosols, which are either washed down by rain or reach the soil through litter fall (Bringmark et al. 1998, Ettler et al. 2005).This explains why forests often contain high Pb concentrations in their topsoils. Due to its toxicity, Pb can have a strong adverse impact on soil microorganisms, and their nutrient cycle functions (Nannipieri et al. 2003). Heavy metal stress generally induces soil microbial communities to employ more energy to maintain metabolic functions and detoxification processes, thereby permitting less energy expenditure for growth. Pb stress has also been reported to reduce plant biomass, respiration rates,and enzymatic activities (Frey et al. 2006).
Heavy metal-contaminated forest soil is recycled using organic matter and native woody plants for remediation. Enzymatic activities are frequently used for determining the influence of heavy metal pollutants on soil microbiological quality (Shen et al. 2005). Heavy metals inhibit enzymatic activity by interacting with the enzyme substrate complexes and denaturing the enzyme protein (Vig et al. 2003). Therefore, soil microbial activities are important to understand the mechanisms and changes in forest ecosystems following metal contamination.
However, total concentrations alone may not convey the entire picture of metal reactivity and its potential toxicity to soil biota. Although the solubility of Pb is generally very low, a considerable fraction of total Pb in soil can exist in soluble forms (Wang and Benoit 1996). The behavior of metals and their availability strictly depends on their chemical form and, thus, on their speciation. Therefore, determining chemical form factors or associations with different mineral phases is of great importance to better understand Pb toxicity in forest soils. An estimate of metal availability is more valuable, as it is related to specific bioavailability, reactivity, mobility, and uptake by plants (McBride 1994). To date, it has generally been accepted that the most appropriate methods for evaluating solid speciation are selective sequential extraction procedures. In fact, a large number of sequential extraction methods have been studied and reported, many of which are variants of the Tessier procedure (Tessier et al. 1979). The mechanisms of metal accumulation in soil lead to five major geochemical forms: (i) exchangeable, (ii) bound to the carbonate phase; (iii) bound to iron-manganese oxides; (iv) bound to organic matter; and (v) a residual metal phase. These metal fractions exhibit remarkable differences in mobility, biological availability and chemical behavior in soil.
Dissolvability, adsorption, and the fraction of heavy metals in soils relate to low molecular weight root exudates, such as organic and amino acids (Kuang et al. 2003). McGrath et al. (1997) reported that Zn bioavailability in soil increases as a result of root exudates. Mench and Fargues (1994) found that the root exudates released from
The pine tree has been widely studied as a bio-indicator for measuring heavy metal toxicity (Poykio and Torvera 2001, Yilmaz 2002). Because pine trees are one of the dominant trees species in eastern Asia, we believe that information on the effects of Pb on pine trees is highly valuable. We report here on the impact of Pb contamination on accumulation and root exudates of
The purpose of this study was to investigate the toxicity of
Natural soil was sampled from the campus of Ewha Woman’s University, which is located in Seoul, Korea. We collected surface soils from a pine tree (
For quality control, a certified reference sample of total metal content in plants was analyzed using the methods described above. Observed concentrations for the certified SRM 1575 (pine needles; National Institute of Standards Technology) were measured. The percentage recovery for the metal of interest was 92.6%.
>
Soil characteristics and enzyme activities
Soil pH was determined by adding soil to water at a ratio of 1:5 (w:v). Soil organic matter content was determined by loss-on-ignition at 700°C in a furnace (MAS 7000; CEM) (Saxena and Bartha, 1983). Soil cation-exchange capacity (CEC) was determined by EPA 9081 methods (US Environmental Protection Agency 1986). Soil nitrate (NO3-) content was determined by extracting soil with deionized water and then measuring NO3- content in the liquid phase using an NO3- electrode (Gelderman and Beegle 1998).
Dehydrogenase activity was measured with an was 2-[4-iodophenyl]-3-[4-nitrophenyl]-5-phenyltetrazolium chloride assay (Tabatabai 1982). The mixtures (fresh soil 3 g) were incubated for 2-h at 37°C. Reaction products were detected using a spectrophotometer (DR/3000 Spectrophotometer; HACH, Loveland, CO, USA) at 485 nm.
The activities of β-glucosidase and acid phosphatase were measured using the methylumbelliferone (MUF)-substrate method (Freeman et al. 1996). The concentration of the MUF-β-glucoside substrate solution was 400 μM (Sigma, St. Louis, MO, USA), whereas the concentration of the MUF-phosphate substrate solution was 800 μM (Sigma). The enzymatic activities in the soil and substrate solution (1:5, w/v) were measured using a fluorometer.
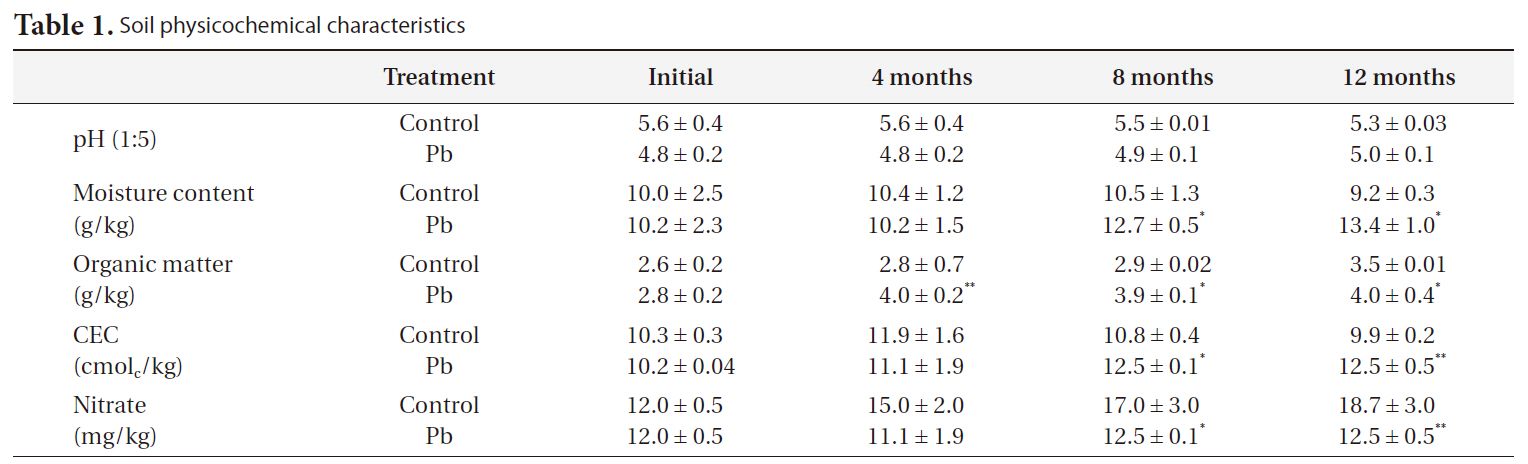
[Table 1.] Soil physicochemical characteristics
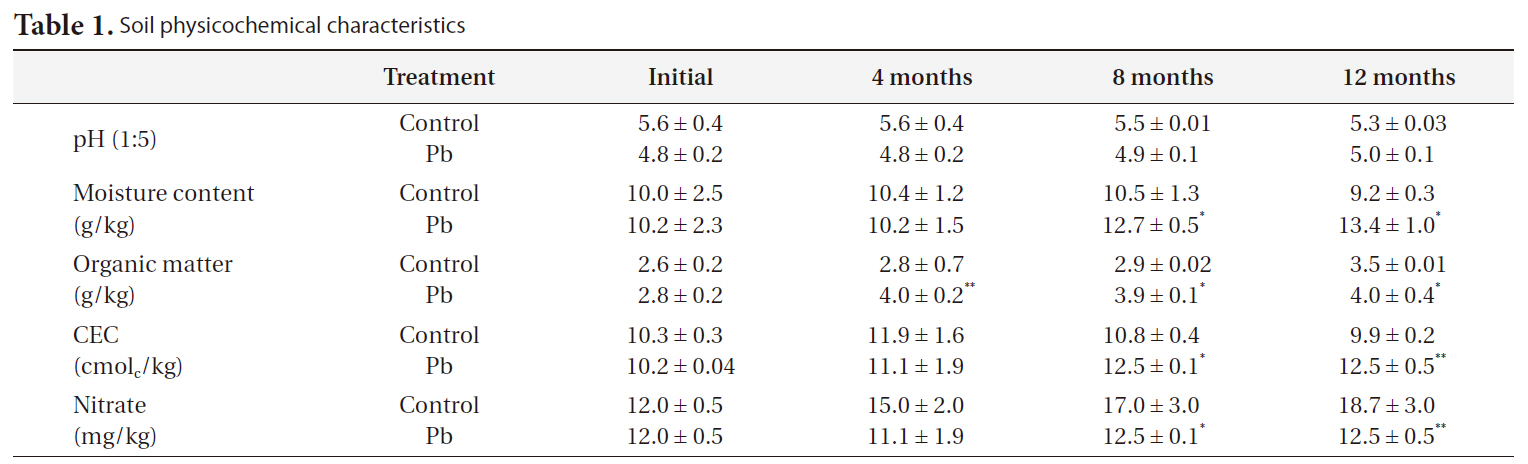
Soil physicochemical characteristics
>
Sequential soil extraction procedures
To determine the amount of Pb in the soils, Pb was fractionated by the sequential extraction procedure of Tessier et al. (1979) in triplicate. Extractions were conducted in 50 mL polypropylene centrifuge tubes. Between each extraction, the supernatant was centrifuged at 6,000 rpm for 15 min and filtered. The five fraction methods indicated below used 1 g of dry soil.
(i) Exchangeable: The soil was extracted at room temperature for 1-h with 8 mL of magnesium chloride solution (1M MgCl2, pH 7.0);
(ii) Bound to the carbonate phase: The residue from (i) was leached at room temperature with 8 mL of 1 M NaOAc adjusted to a pH of 5.0 with acetic acid (HOAc). Continuous agitation was maintained, and the time necessary for complete extraction was evaluated;
(iii) Bound to iron-manganese oxides: The residue from (ii) was extracted with 20 mL 0.4 M NH2OH HCl in 25% (v/v) HOAc. The latter experiments were performed at 96 ± 3°C with occasional agitation, and the time needed for complete dissolution of the free iron oxides was evaluated;
(iv) Bound to organic matter: To the residue from (iii), 3 mL of 0.02M HNO3 and 5 mL of 30% H2O2 were added and adjusted to a pH of 2 with HNO3, and the mixture was heated to 85 ± 2°C for 2-h with occasional agitation.
A second 3 mL aliquot of 30% (v) residual metal phase: The residue from (iv) was digested with a HF-HClO4 mixture according to the procedure described below for total metal analysis.
(vi) Total: Fresh sample (1.0 g) was digested with 8 mL of aqua regia (HCl and HNO3, 3 + 1, v/v) for 2-h at 120ºC.
To verify the reliability of our sequential extraction procedures, recovery (%) of the sum of the Pb concentrations in individual fractions to the total Pb concentration in soils was calculated and found to be 102.5%.
Plants were harvested by gently removing them from the soil. Prior to analysis, the plants were washed with water to remove soil particles. The plants were sampled and the extracts were prepared as described by Yun and Kil (1992). Fresh roots (100 g) were immersed in 1 L of distilled water for 48-h, after which the solutions were filtered through a 0.45 ㎛ syringe filter and used undiluted.
Organic acids were analyzed with an ion chromatography unit (DX-500; Dionex, Sunnyvale, CA, USA) equipped with an ED40 electro-chemical detector, an IonPac AS11 (4 × 250 mm) column, and an ASRS-Ultra II Anion self-regenerating suppressor (CA). The total concentration of the root organic acids was calculated as the sum of the concentrations of individual organic acids.
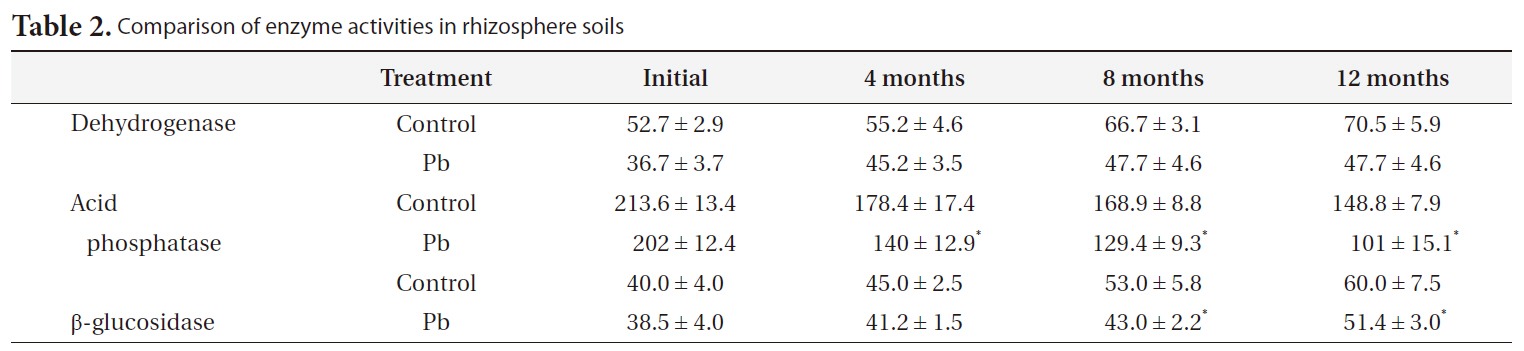
[Table 2.] Comparison of enzyme activities in rhizosphere soils
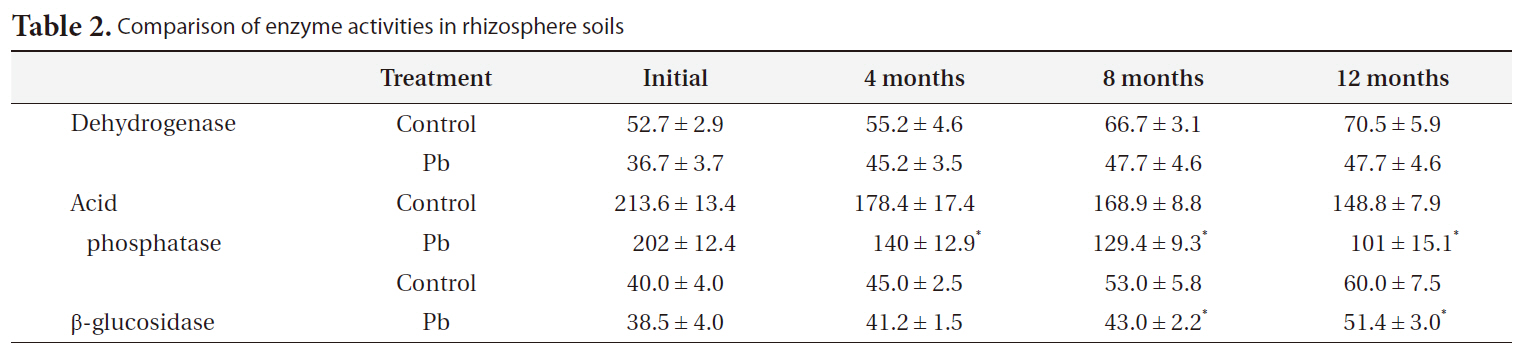
Comparison of enzyme activities in rhizosphere soils
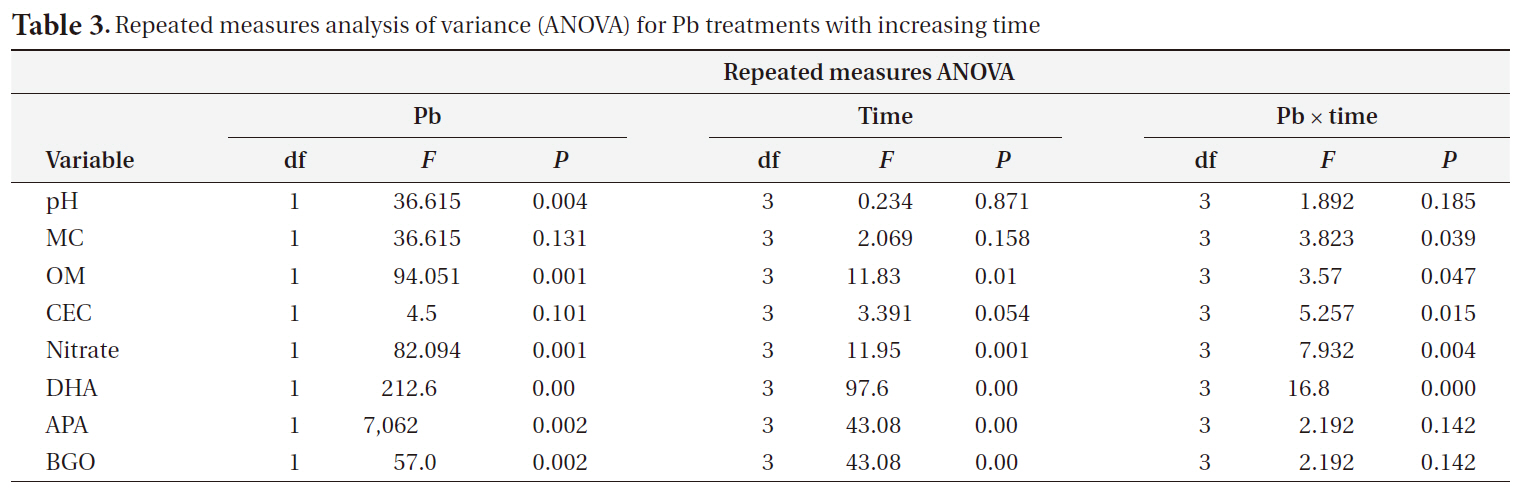
[Table 3.] Repeated measures analysis of variance (ANOVA) for Pb treatments with increasing time
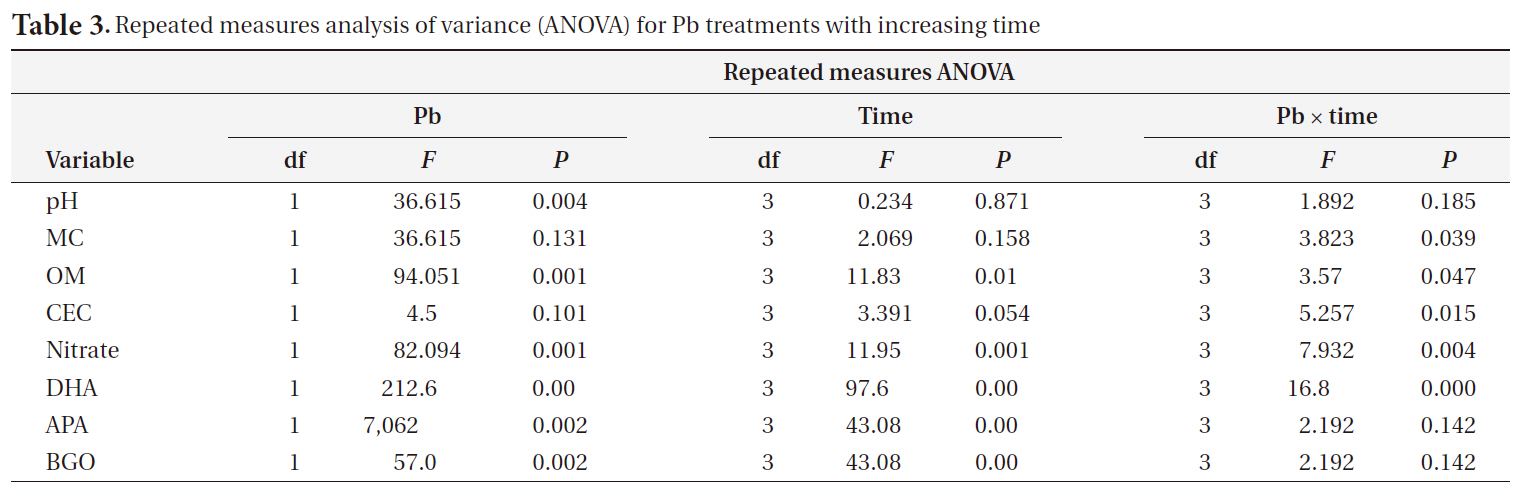
Repeated measures analysis of variance (ANOVA) for Pb treatments with increasing time
We used a repeated-measures analysis of variance (ANOVA) to determine the effects of time and Pb treatment on the physical and chemical characteristics of the soil and on soil enzyme activity using SPSS ver. 17.0 (SPSS Inc., Chicago, IL, USA). The relationship between metal uptake and the metal fraction was determined using a classic multivariate linear regression model on Microsoft Excel.
>
Soil characteristics and enzyme activity
The soil characteristics are shown in Table 1. The physicochemical properties of the soil were as follows: Soil texture, loamy sand (54.1% sand, 30.9% silt, and 15.0% clay); total organic matter, 2.8%; total moisture content, 10%; CEC, 10.3 mmol/kg. Soil pH was in the weakly acidic range (5.6-6.2).
Table 2 describes the soil enzyme activity. Dehydrogenase, acid phosphatase, and β-glucosidase decreased remarkably with increasing Pb (P < 0.01) (Table 2). Relative to the controls, dehydrogenase, acid phosphatase, and β-glucosidase activity in the Pb treatment group decreased 33%, 32%, and 12%, respectively. However, in Pb-contaminated soils, dehydrogenase and β-glucosidase increased with time. In contrast, acid phosphatase activity decreased slightly over the same time period.
The pH decreased slightly in Pb-contaminated soils due to the use of the PbSO4 solution, although it did not significantly change over time. Organic matter and nitrate were greatly influenced by both Pb and time. Additionally, there were Pb-X time interactions, as indicated by the repeated-measures ANOVA (Table 3). Organic matter increased with time and Pb treatment. Nitrate decreased with Pb treatment but increased with time.
>
Changes in the soil Pb fraction and uptake by Pinus densiflora
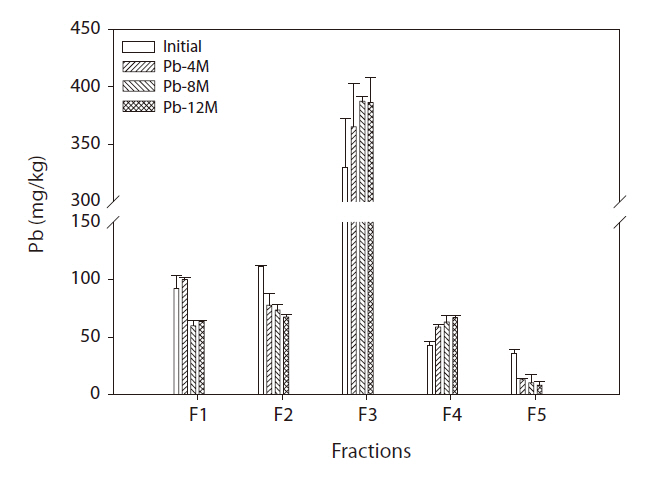
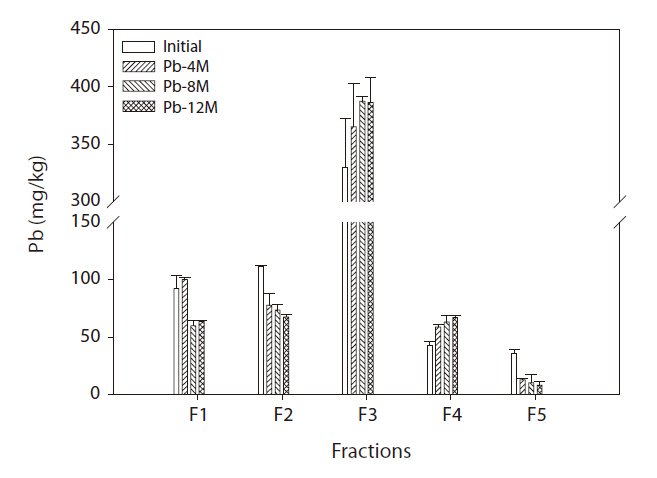
The soil Pb binding forms are presented in Fig. 1. Most of the Pb soil fraction was bound to iron-manganese oxides. The percentage present as exchangeable (F1) and Pb-bound to carbonate (F2) fractions and bioavailability decreased over time, although the F3 and F4 fractions increased over time. The amount of Pb bound to Fe-Mn oxides (F3) and bound to organic matter (F4) increased consistently with increasing incubation time. After 8 months of incubation, the Pb concentration in the Fe-Mn oxide-bound fraction in soil reached 358 mg/kg. Pb concentrations were 62 mg/kg in the organic matter fraction.
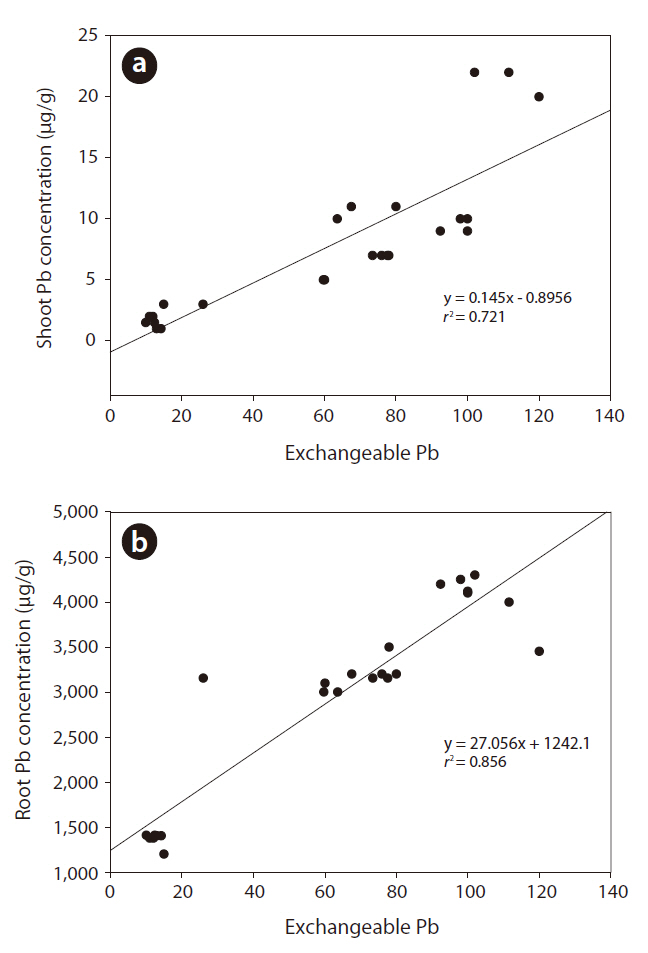
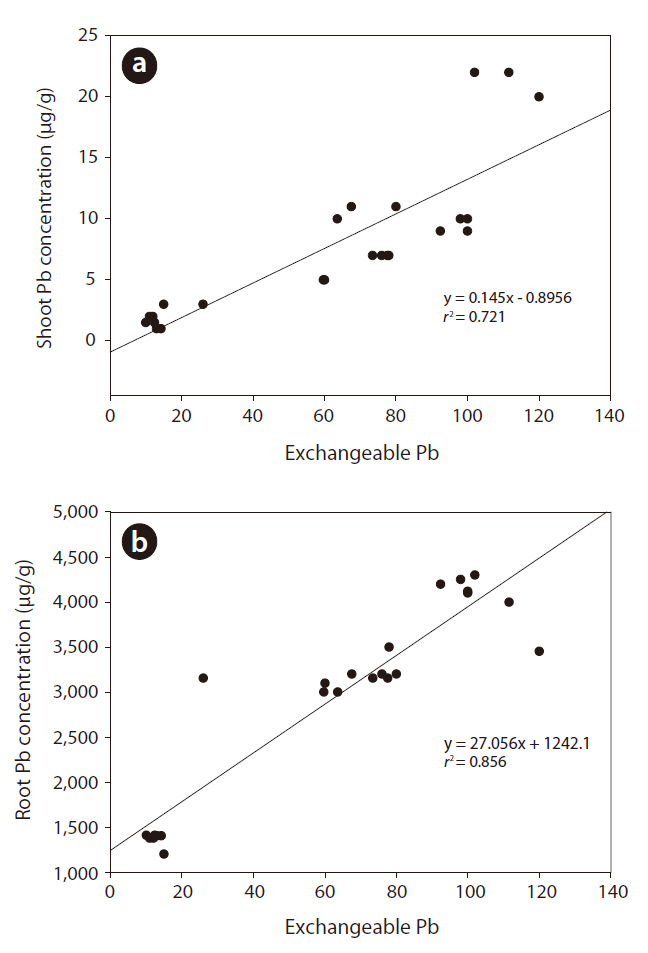
Fig. 2 shows the correlations between soil exchangeable Pb concentrations and pine tree uptake of Pb. Pb uptake by roots and shoots was positively correlated to an increase in exchangeable Pb in the soil. Absorption and accumulation of Pb in the roots and shoots increased with time. The Pb content bound to roots increased to nearly 90% upon exposure to Pb.
>
Organic acids in root exudates
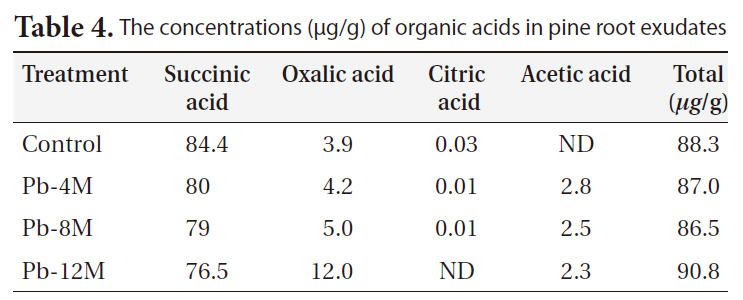
Table 4 shows the organic acid concentrations of pine root exudates, which affected growth rate and heavy metal uptake. The primary organic acid in the pine exudates was succinic acid. Furthermore, small amounts of oxalic acid, acetic acid, and citric acid were detected in root exudates. Specifically, the amount of oxalic acid was 4-fold greater in the roots of pine in the pots treated with Pb after 12 months, although succinic acid decreased during the incubation period. Acetic acid was only detected in
[Table 4.] The concentrations (μg/g) of organic acids in pine root exudates
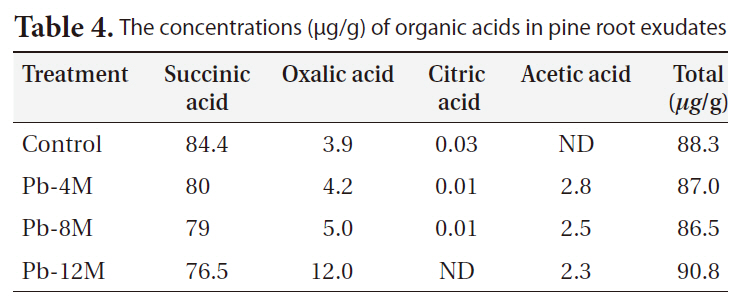
The concentrations (μg/g) of organic acids in pine root exudates
Pb-contaminated soils.
>
Changes in soil characteristics and enzyme activity
Enzyme activity significantly decreased in Pb-contaminated soils. Such a decrease in microbial activity was concomitant with a marked increase in exchangeable and soluble Pb. Effects exerted on microbial activity may be attained by lowering the soil pH with organic acid (Shotyk et al. 1998), and complexation with heavy metals (Wierzbicka 1999), clay minerals and metal oxides with organic ligands from bacterial exudation (Gadd 2000). The effects of soil pollution on enzyme activity are complex; the response of different enzymes to the same pollutant may vary greatly, and the same enzyme may respond differently to different pollutants (He et al. 2003). In this study, Pb inhibited all enzyme activity, whereas Pb only slightly inhibited organic C-acquiring enzyme activity (β-glucosidase). It has been reported that different metal ions exhibit different behavior in their ability to act as β -glucosidase inhibitors (Scigelova and Crout 1999).
However, dehydrogenase and β-glucosidase increased over time in the rhizosphere. Under the influence of root activity, Pb extracted from polluted soils increased, as pH decreased. Exchangeable Pb was much higher in the rhizosphere than that in bulk soil. Plants can modify metal speciation and behavior in the rhizosphere by producing exudates (Laperche et al. 1997), and these results were time dependent. The pine exudates may have been affected by an interaction with microbial activities and soil properties in the rhizosphere. Some researchers have studied the relationships between the pine rhizosphere and metal toxicity. Hartley-Whitaker et al. (2000) reported that Cd and Zn accumulation in
>
Correlation between soil exchangeable Pb and shoot and root Pb concentrations
Fig. 1 shows the time-dependent changes in Pb concentration of each fraction. Cultivation time significantly affected the Pb fraction distribution. The newly added Pb existed mainly in the surface soil particles as an exchangeable fraction.
Fig. 2 presents the correlations between soil exchangeable Pb concentrations and pine tree uptake of Pb. Exchangeable metals in the soil are easily available for plant uptake (Kabata-Pendias 1993). Metal toxicity and accumulation in plants are related to the effects of synergetic and antagonistic metal interactions. These effects may vary depending on the physicochemical factors such as organic matter content, pH, redox potential, and ion speciation. These factors could influence metal mobility and bioavailability in soils (van Gestel 2008). For example, lower soil pH favors the release of metals into soil solution, allowing plants to take up more metals (Tang et al. 2003, Wu et al. 2009). In the present study, soil pH decreased slightly with the addition of Pb (Table 1). Moreover, a high organic matter content in soil resulted in increased organic matter in the bound fractions (F3 and F4) over time (Table 2). These results show that the metal fraction in soil was related to changes in the organic matter content due to microbial activity (Lock and Janssen 2001). Investigations relative to these questions should complement the present approach for a full understanding of the multifaceted issue of bioavailability in soil systems.
>
Organic acids in root exudates
An increase in Pb uptake was associated with increases in root exudates, suggesting that the root exudates activate Pb by dissolution and chelation. Because pH is a key factor controlling heavy metal extractability and mobility, changes in organic acids in root exudates may modify Pb availability by changing the pH (Zhang et al. 2006). Other studies have shown the significance of root exudates in the bioavailability of heavy metals in soil (Chen et al. 2006, Zhang et al. 2006). High molecular weight organic acids, such as humic acid, can reduce the bioavailability and toxicity of heavy metals (Liao and Huang 2002), whereas low molecular weight organic acids and amino acids can increase the bioavailability and plant accumulation of heavy metals by decreasing the rhizosphere pH or chelating metals in the soil (Liao and Huang 2002).
The results of this study demonstrate that changes in the soil metal fraction are related to soil chemical characteristics, microbial activity, and root exudates. The exchangeable metal content increased in soil with a low pH and the presence of organic acids from root exudates. In addition, root exudates change microbial activity that use the main C-sources: d-malic acid, d-glucosaminic acid, and α-ketobutyric acid (data not shown). These findings indicate that organic acids from plant roots can combine with metal ions, which can then be taken up easily by microbes and plants.