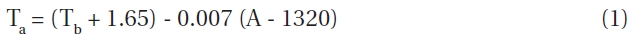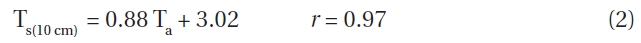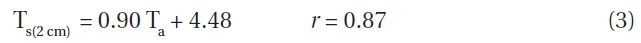Studies on the vertical segregation of plants in Japan were initiated by Imanishi (1949) in the North Alps of central Japan. Thereafter, Takahashi (1962) described the vertical distribution of mountain forests in Central Honshu, and more recently Ohsawa (1984) examined the distribution of forest communities at different elevations on Mt. Fuji. These studies mainly focused on the floristic composition and structure of the forests. Betula platyphylla var. japonica (Miq.) Hara and Betula ermanii Cham. segregate vertically at an elevation of 1,850 m on the southern slope of Mt. Neko in Nagano Prefecture, Japan. The altitudinal change in floristic composition on Mt. Neko was studied first by Tanouchi and Hayashi (1981), but no causative relationship between environmental parameters and the altitudinal distribution of B. platyphylla var. japonica and B. ermanii was discussed. We have attempted to clarify the factors affecting the vertical distribution of these species by relating ecological traits such as leaf phenology and seed germination to environmental conditions on the mountain (Kikuzawa 1983, Kudo 1995, Chuine and Beaubien 2001). In this study, those ecological characteristics affecting to the thermal environment of these species were examined through germination, leaf phenology, and the growth of seedlings (Raulier and Bernier 2000)
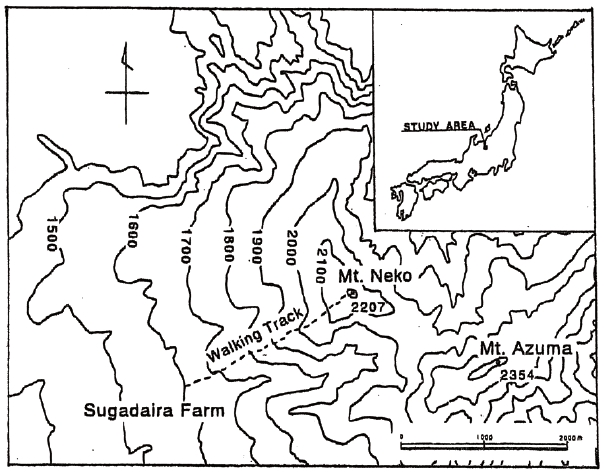
Sugadaira (36o31' N, 138o21' E), located at an elevation of 1,320 m (Fig. 1), has a mean annual air temperature and total annual precipitation of 6.5℃ and 1,102 mm, respectively. The area lies in the cool temperate zone and supports a climax forest of summer-green, broad-leaved trees dominated by Fagus crenata Blume. Above around 1,850 m, these species are replaced by evergreen conifers such as Abies veitchii Lindl. Most of the area in Sugadaira is occupied by secondary forest consisting of Quercus crispula Blume, Pinus densiflora Sieb. et Zucc., B. platyphylla var. japonica and B. ermanii interspersed with plantations of Larix leptolepis Sieb. et Zucc. Gordon, farms, ski slopes and cultivated lands (Tanouchi and Hayashi 1981). The soils are Andosols, which have developed on volcanic deposits from Mt. Azuma and Mt. Neko (2,207 m) slopes gently to the south with steeper slopes on its northern flank, which are associated with the old crater. Distinct forest zones can be observed at different elevations on the mountain, with P. densiflora being dominant at 1,300-1,500 m. At 1,500-1,800 m, the forests are mainly composed of B. platyphylla var. japonica which is then replaced on the upper slopes by B. ermanii (1,800-2,100 m). A. veitchii is the dominant species at 2,000-2,100 m. Tanouchi and Hayashi (1981) suggested that this zonation reflected the interaction between the forests and environmental conditions and human activities. The study site was located on the southern slope of Mt. Neko, extending from 1,600 m to the summit.
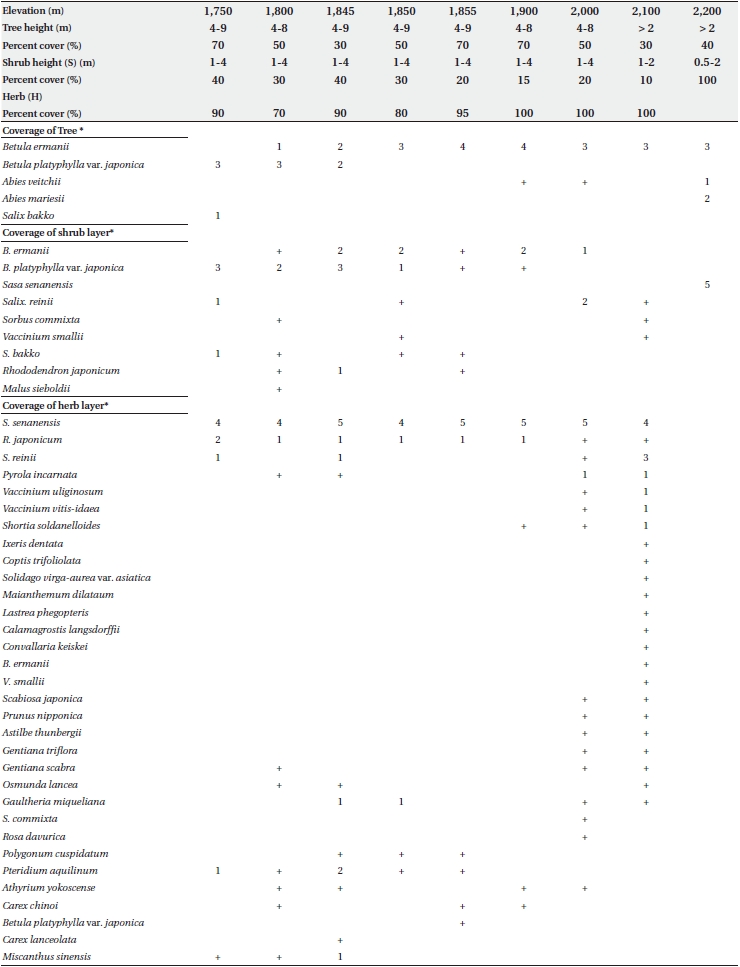
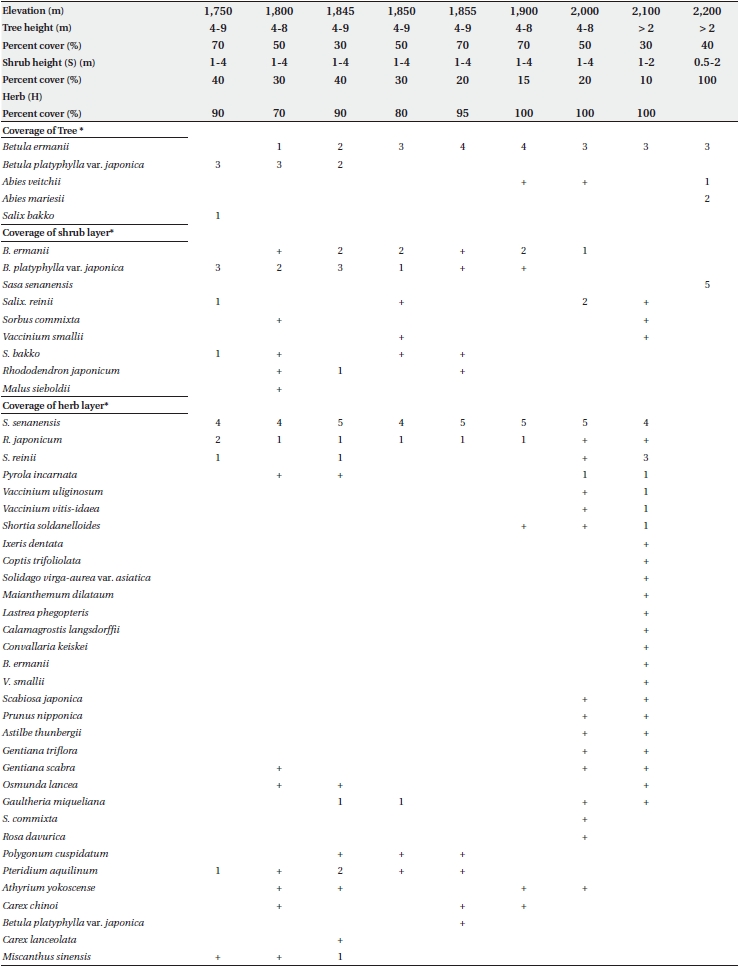
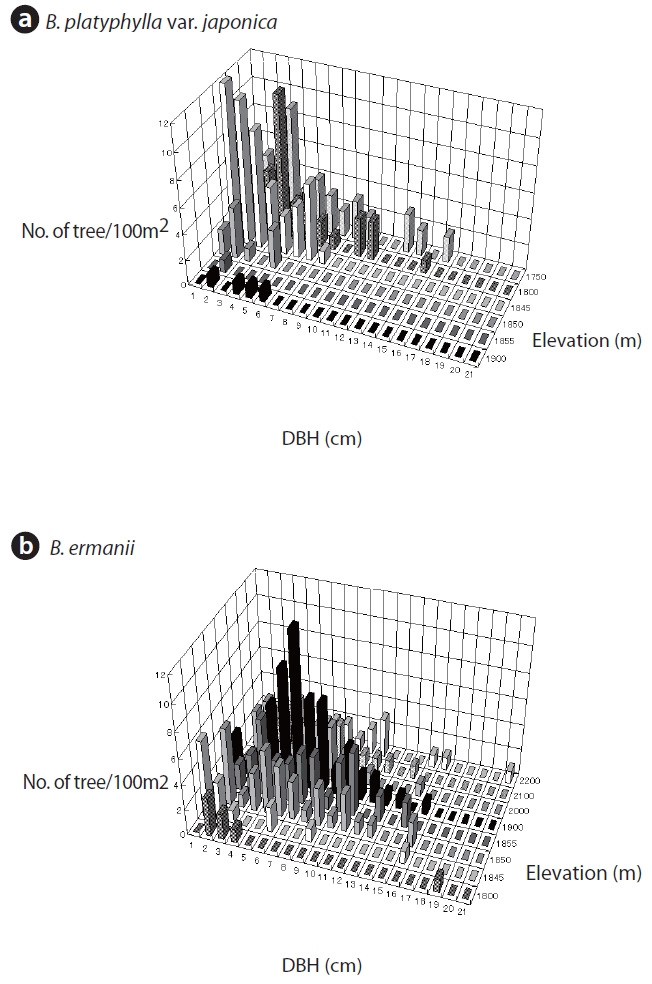
Floristic composition, trunk diameter at breast height (DBH) for trees larger than 1 cm diameter, and tree height was assessed in 10 × 10 m quadrats located at elevations of 1,750, 1,800, 1,850, 1,900, 2,000, 2,100, and 2,200 m. Additional quadrats were sampled at 1,845 m and 1,855 m, in the transitional zone between B. platyphylla var. japonica and B. ermanii. The trunks of both B. platyphylla var. japonica and B. ermanii often sprouted from stem bases, and in this survey, such sprouts were considered to be single trees. Field work was conducted from 8-20 September 1995.
Auto-recording thermometers (Corner System LcII; Corner System Co Ltd. Sapporo, Japan) were used to measure air and soil temperatures. These were placed in an open area on the summit of Mt. Neko (2,200 m) and at elevations of 1,700 m (mid-elevation of the B. platyphylla var. japonica zone), 1,850 m (boundary between the B. platyphylla var japonica and B. ermanii zones) 1,960 m (mid-elevation of the B. ermanii zone). Above-ground air temperature (1.3 m) and soil temperature (10 cm depth) were recorded at hourly intervals from June 1995 to August 1997. During the last 10 days of August 1997, soil temperature at a depth 2 cm was also measured at 1,850 m.
Germination tests were conducted on B. platyphylla var. japonica and B. ermanii seeds collected at elevations of 1,750 m (28 September 1995) and 2,050 m (18 September 1995). The seeds of both species were incubated under constant temperature conditions at 10, 15, 20, 25, and 30℃, and under alternating temperature conditions at 10/20, 15/25, and 10/30℃ (16 hours in low temperature, 8 hours in high temperature). Germination was conducted under a 16L:8D regime for 40 days, with the light intensity in the chamber being 80 mol m-2 s-1 for the duration of the experiment. Forty seeds were sown in each Petri dish with five replicates performed for both species and seedling emergence was monitored daily. In addition, we examined the germination of seeds which stratified at 4℃ for 134 days.
Twelve 50-day-old B. platyphylla var. japonica seedlings and four B. ermanii seedlings were planted at uniform distances in a deep plastic pot (10 cm ×φ9 cm) with four replications. A mixture of a typical Andosol and immature soil (Akadama-tsuchi) was used for cultivation. The seedlings were grown under temperature conditions of 10, 14, 20, and 30℃ under constant light (60 mol m-2 s-1) for 40 days. Seedlings were harvested on days of 21, 47, 67, and 90 after planting and survival rates were calculated before weighing after drying in an oven for 2 days at 70℃. In order to assess seedling establishment under field conditions, we transplanted ten 30-day-old seedlings of each species to elevations of 1,600, 1,850, 2,050, and 2,200 m on 21 July 1997. The seedlings were planted in unglazed pots (φ14 cm × 14 cm) with the same soil as the that used in the laboratory experiments. We recorded seedling survival for each pot at each altitude on 29 August and 8 October, which were 52 and 92 days after planting, respectively.
Leaf unfolding in B. platyphylla var. japonica and B. ermanii trees growing in close proximity was related to a modified Kira's warmth index (WI) (Kojima et al. 2003), which was calculated as the sum of the mean daily temperatures above 5℃ expressed as ℃ day. The annual WI was calculated by summing the mean daily temperature from January 1 for each year. The stages of leaf unfolding identified were as follows (Kojima et al. 2003):
L0: winter bud.
L1: a bud swells and its end splits so that green tissues are exposed.
L2: distal end of leaf open and leaf unfolding has begun.
L3: joint of first leaves opens and a clear leaf shape can be identified.
L4: second leaves begin unfolding.
L5: all leaves completely unfolded and maximum leaf area attained.
RESULTS
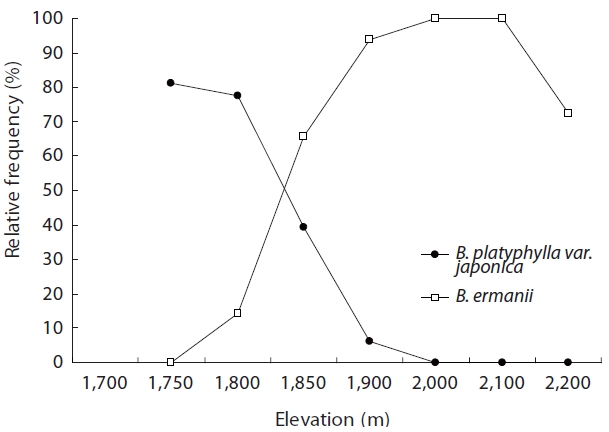
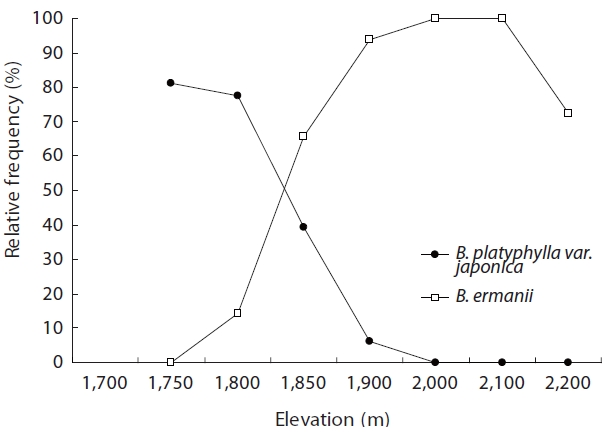
Table 1 shows the floristic composition of selected elevations on Mt. Neko. Species dominance of trees changed from B. platyphylla var. japonica to B. ermanii at 1,850 m. The shrub stratum at higher elevations was comprised mainly of Vaccinium vitis-idaea Linn. and Salix reinii Franch. et Savatit while Rhododendron japonicum (A Gray) Suringer, Sasa senanensis Franchet et Savatier Rehder var. senanensis and Miscanthus sinensis Anderss were common at lower elevations. At higher elevations, Vaccinium uliginosum Linn was abundant in the shrub layer. S. senanensis was the most dominant herbaceous species at all elevations. Except for the tree species, no marked changes in floristic composition were observed at 1,850 m in the transitional zone between the B. platyphylla var. japonica and B. ermanii communities. The change in the relative frequency of B. platyphylla var. japonica and B. ermanii and the abrupt transition in dominance at 1,850 m are shown in Fig. 2. From 2,000 to 2,100 m B. ermanii constituted the canopy tree species, but at the summit it grew in association with A. veitchii and A. mariesii Masters. At around 1,750 m, Salix bakko Kimura and Salix commixta Hedl. appeared in the understory, resulting in a slight decrease in the relative frequency of B. platyphylla var. japonica.
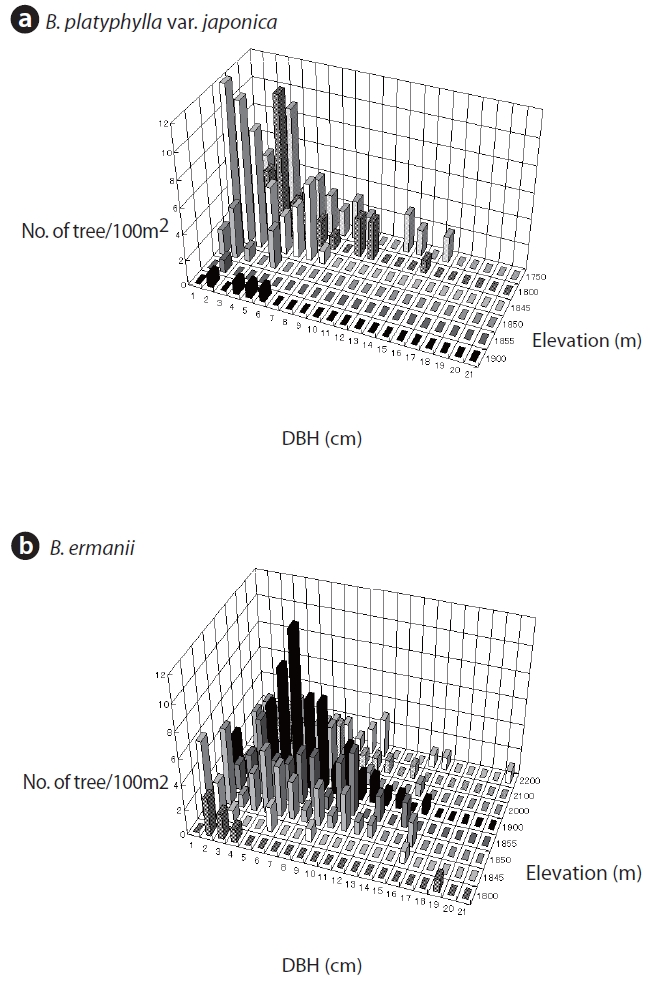
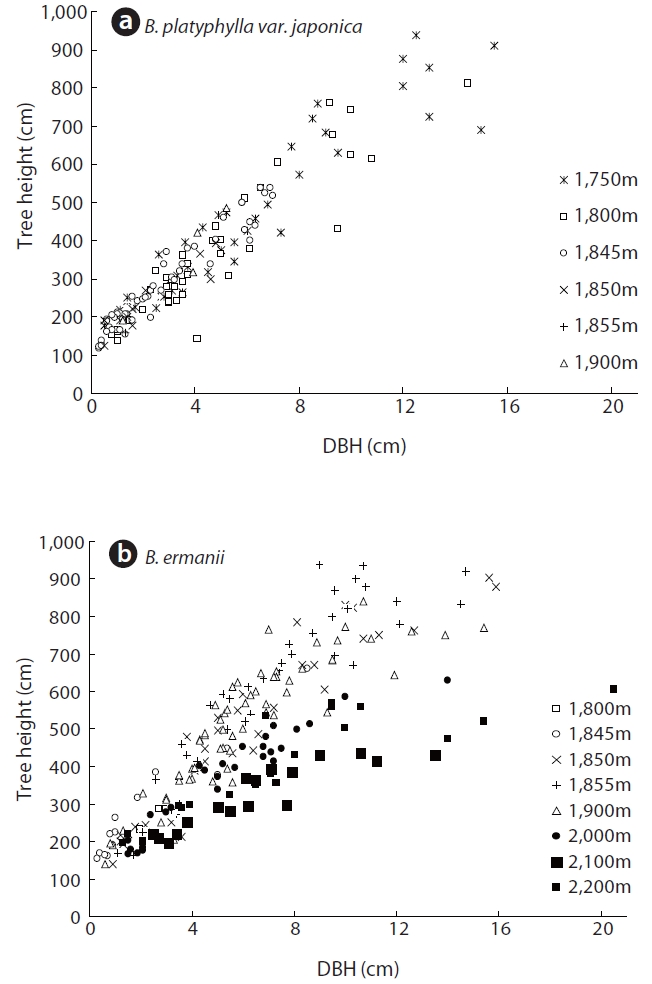
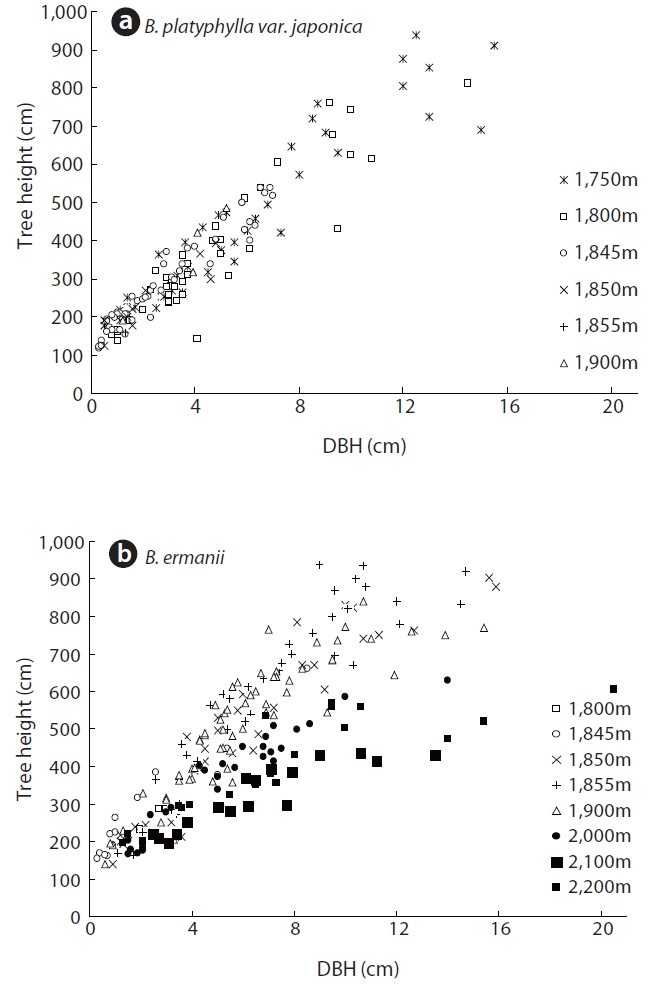
The changes in DBH and tree density for B. platyphylla var. japonica and B. ermanii with elevation are shown in Fig. 3. The DBH of most trees ranged between 1 cm and 12 cm, with the density of B. platyphylla var. japonica decreasing with elevation while that of B. ermanii increased. The trees of B. platyphylla var. japonica have not distributed in the site above 2,000 m. The change in DBH versus tree height at different elevations for B. ermanii and B. platyphylla var. japonica is presented in Fig. 4. The ratio of tree height to DBH was more variable in B. ermanii than B. platyphylla var. japonica, particularly for larger trees. Thus, B. ermanii trees with DBHs exceeding 12 cm ranged between about 4 m to 9.5 m in tree height compared to about 7 m to 9.5 m for B. platyphylla var. japonica. The plasticity in tree growth form of B. ermanii was greater than that of B. platyphylla var. japonica. At the elevation above 2,100 m, the ratio of tree height to stem diameter of B. ermanii showed the small value markedly compare to the trees inhabiting in the lower elevations. This suggests an adaptive trait for mountain environments including strong wind.
The rate of sprouting from stumps was 17-80% for B. ermanii compared to 33-61% for B. platyphylla var. japonica, and this was not significant (P < 0.05).
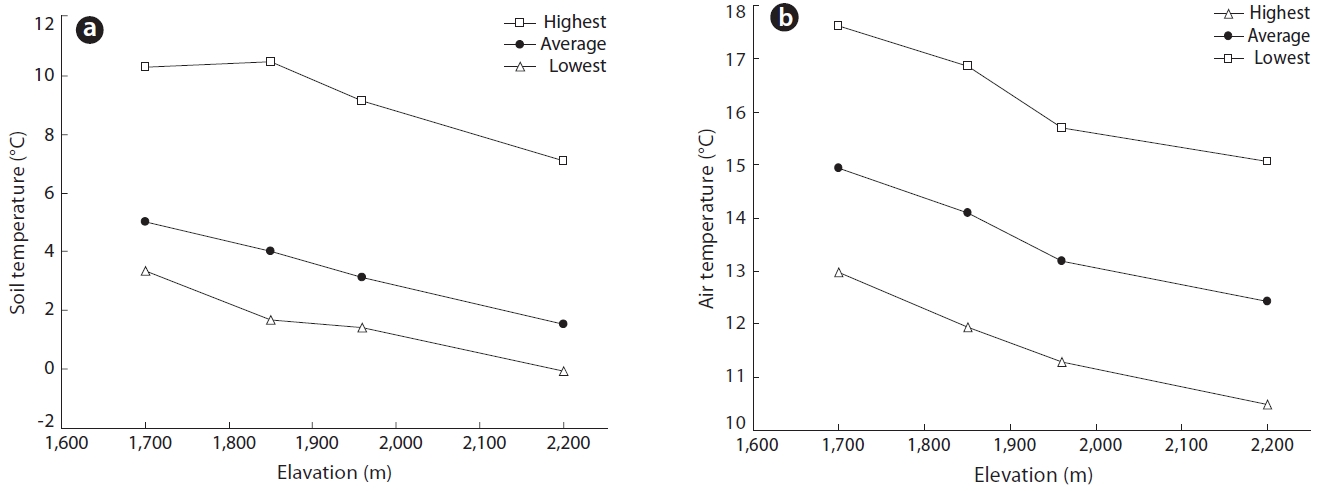
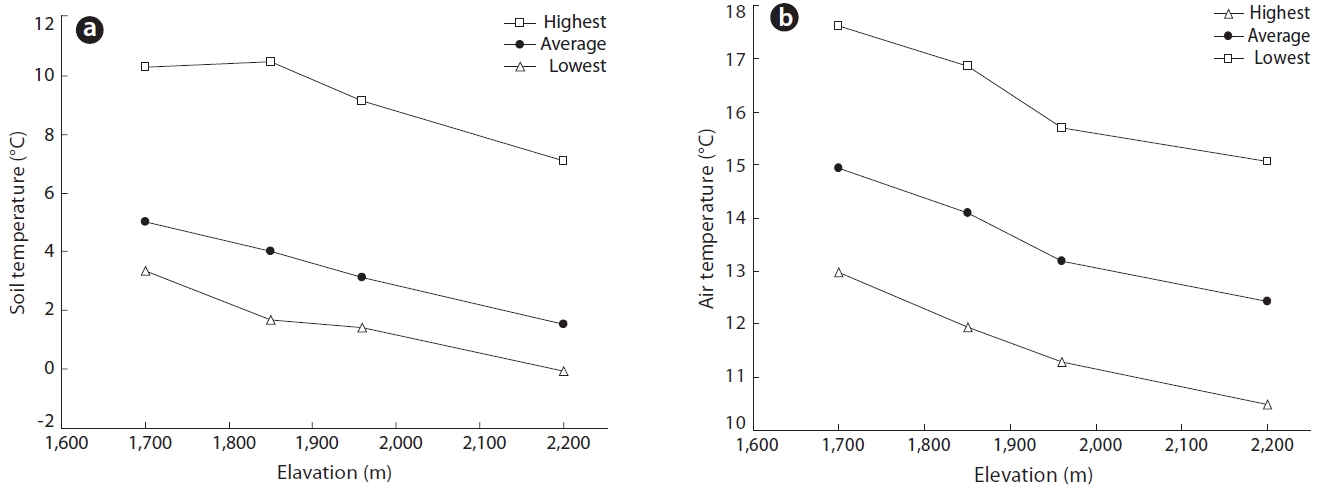
The mean annual temperatures for each elevation were 5.0℃ in 1,700 m, 4.0℃ in 1,850 m, 3.1℃ in 1,950 m and 1.5℃ in 2,200 m. The annual average, average highest and average lowest temperatures, as well as the soil temperatures at a depth of 10 cm in growing season from April to September over the elevation range 1,700 to 2,200 m are shown in Fig. 5. Mean air temperature declined linearly with elevation at a lapse rate of 0.7℃ per 100 m. In 1996, the mean air temperature during growing season at 1,850 m was 14.1℃
The relationship between mean annual temperature at a given elevation on Mt. Neko and at the Sugadaira Montane Research Centre at Tsukuba University (SMRCT) at 1,320 m was estimated by the following equation:
Given the strong correlation between the temperatures at both sites (r = 0.98) this equation was used to estimate the mean air temperature at any elevation using the measured values obtained at SMRCT. Using this method, the mean annual temperature at 1,850 m was estimated to be 4℃ and 14.1℃ during growing season between 1991 and 1996. These findings closely corroborated the values measured in 1996. Average annual minimum and maximum air temperatures also decreased with elevation, being 10℃ and 20℃, respectively.
Mean air temperature (Ta) and mean soil temperature (Ts) at depths of both 2 cm and 10 cm were strongly correlated at all elevations. For example, at 1,850 m, soil and air temperatures could be represented as follows:
Similar correlation coefficients were obtained for all elevations. We used equation (3) to elucidate which soil temperature conditions where important for seed germination. At 1,850 m, the difference between the mean daily air and soil temperatures at a depth of 2 cm were ≥ 10℃ for 25 days, ≥ 15℃ for 17 days and ≥ 20℃ for 5 days for during May to October 1996. The average daily difference in soil temperature was 15℃ at 1,850 m.
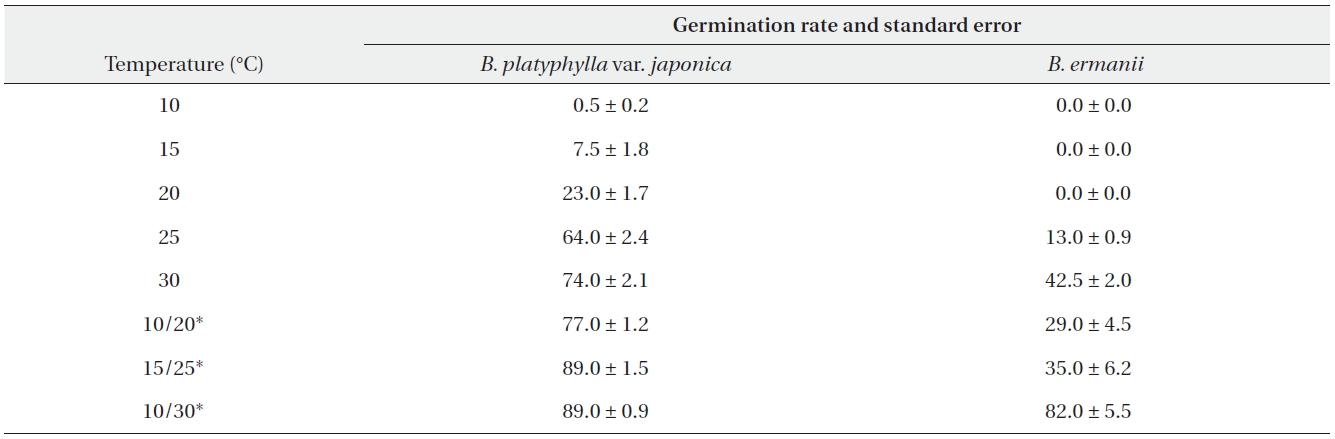
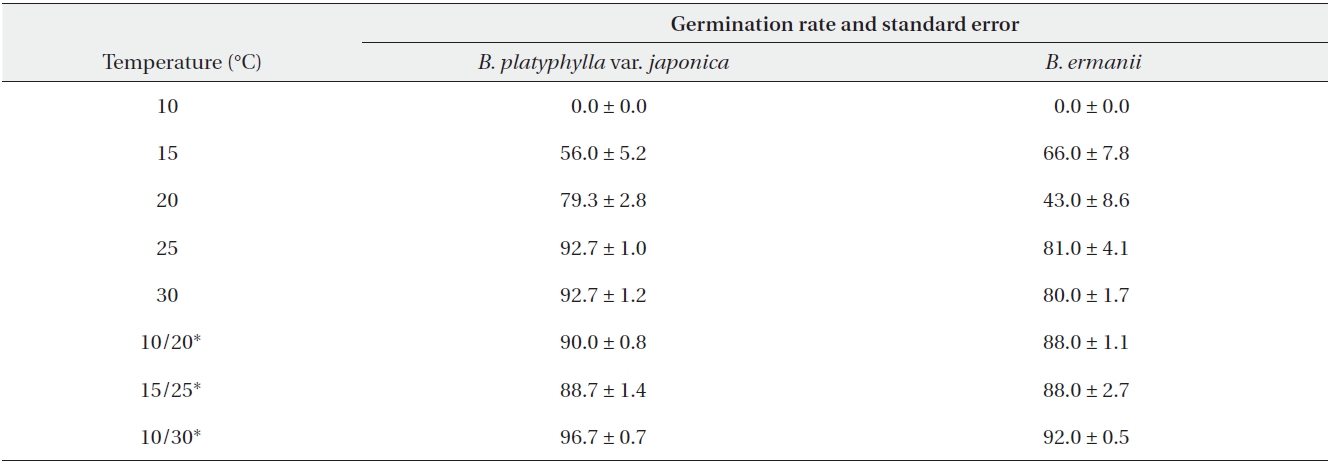
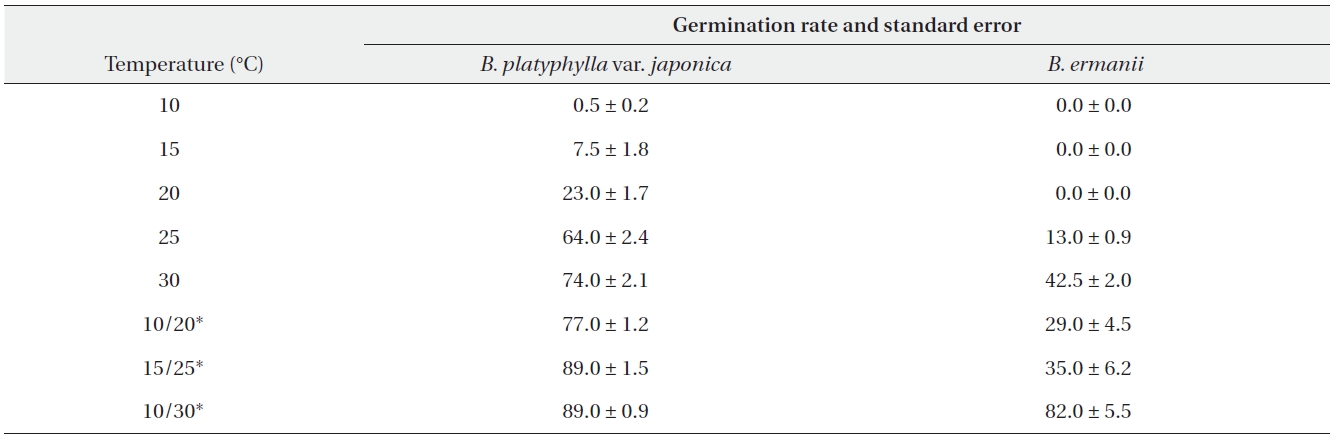
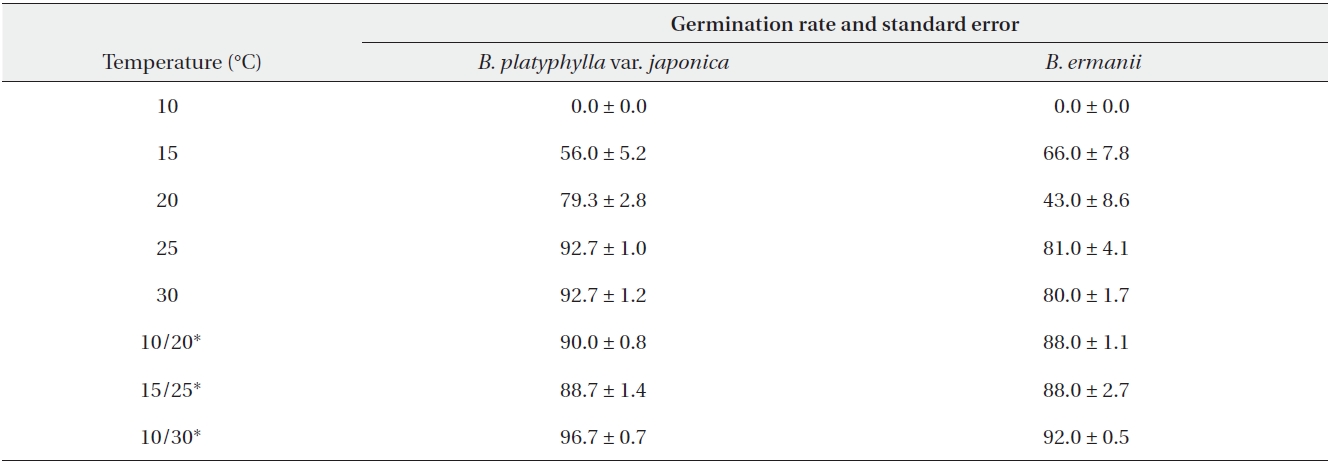
The germination rates of B. platyphylla var. japonica and B. ermanii under different temperature regimes are given in Table 2. For both species, the maximum germination rate at a constant temperature was observed at 30℃ when 74.0% and 42.5% of B. platyphylla var. japonica and B. ermanii germinated, respectively. At a constant temperature of 10℃, only 0.5% of the B. platyphylla var. japonica germinated. No seeds of B. ermanii germinated below 20℃. Indeed, under the same temperature conditions, germination B. platyphylla var. japonica was consistently higher than that observed in B. ermanii. Under the condition of alternating temperature, 77-89% of B. platyphylla var. japonica germinated. For B. ermanii, 82% of seedlings germinated under the 10-30℃ regime. The germination rates after cold stratification at 4℃ are shown in Table 3. Stratification greatly promoted the germination in both species, but was particularly effective for B. ermanii in which 66% and 43% of seedlings germinated under constant temperatures of 15℃ and 20℃, respectively.
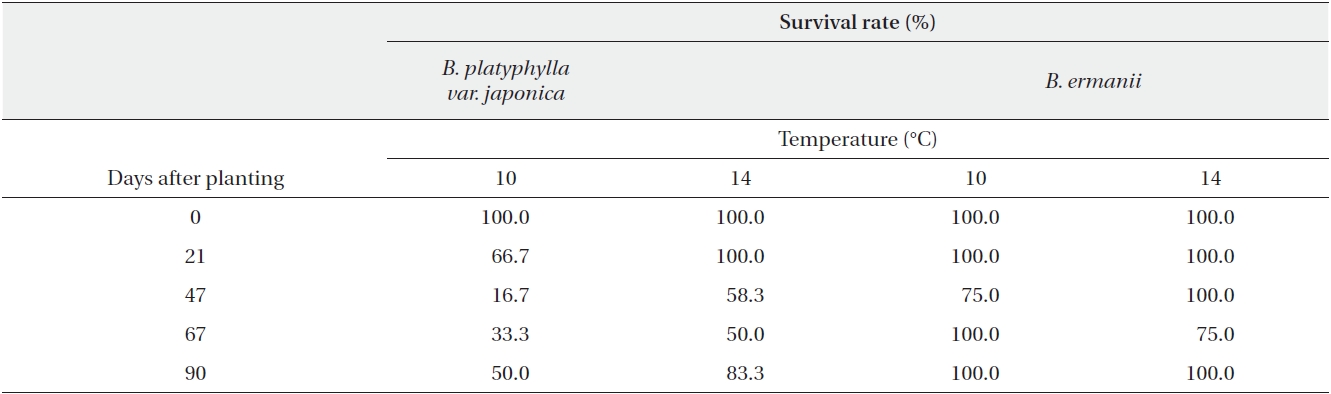
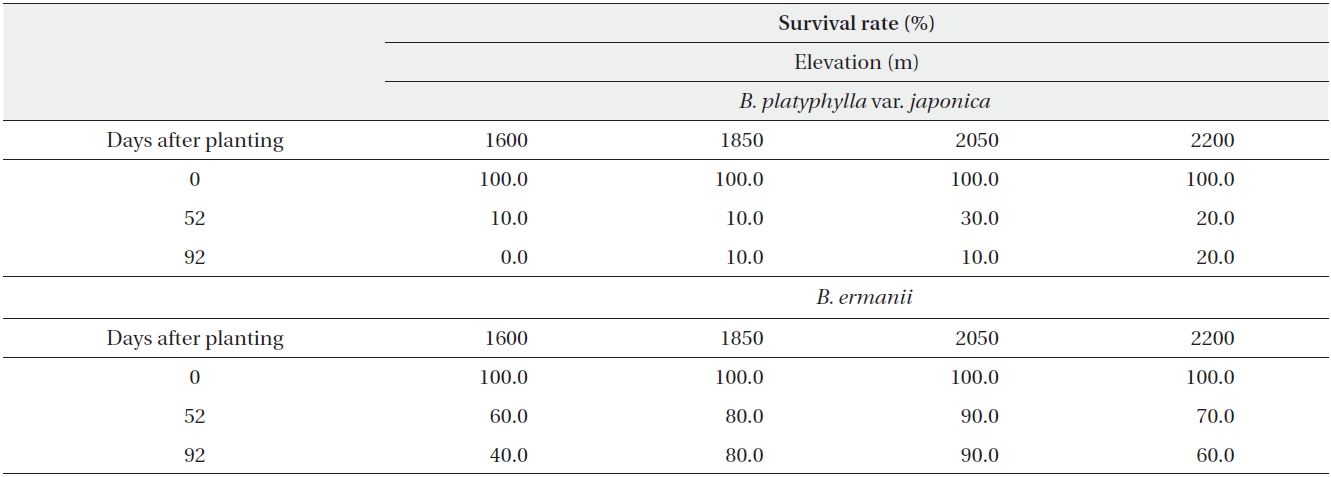
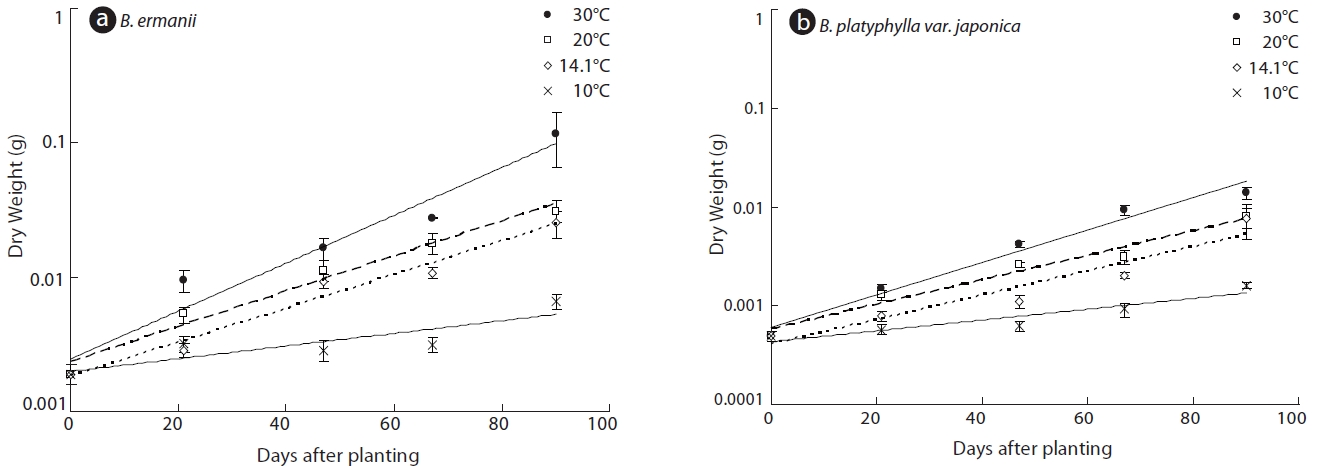
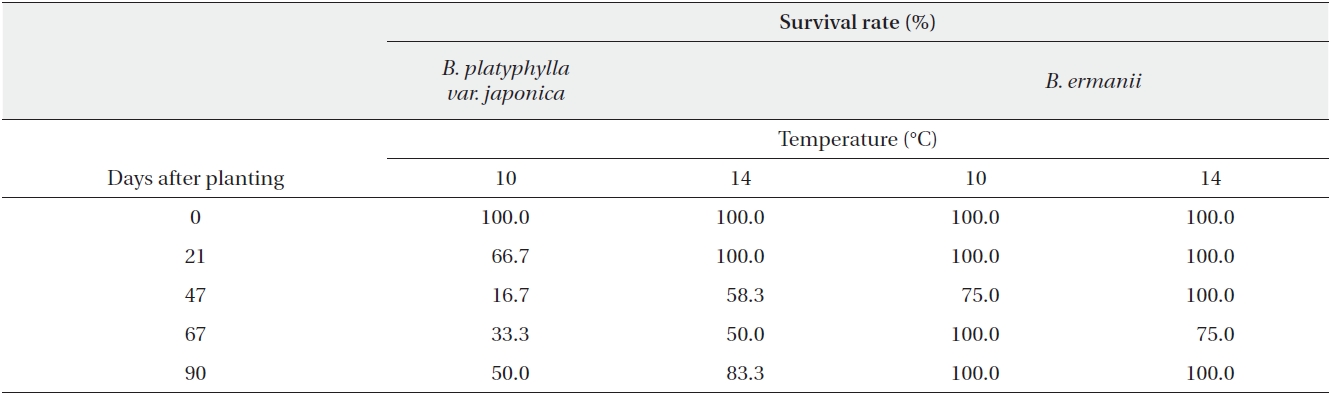
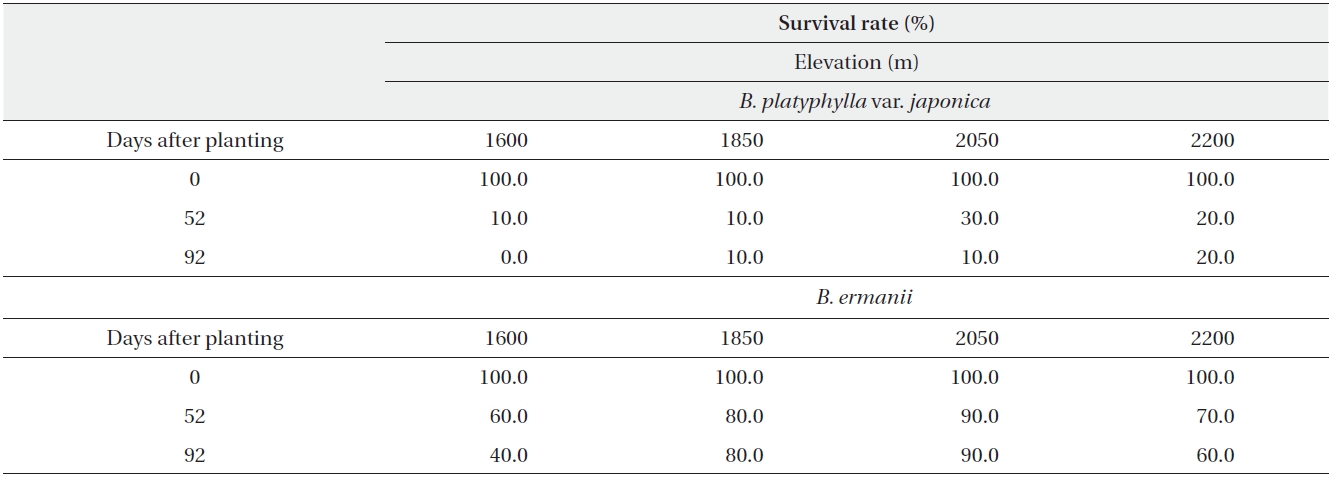
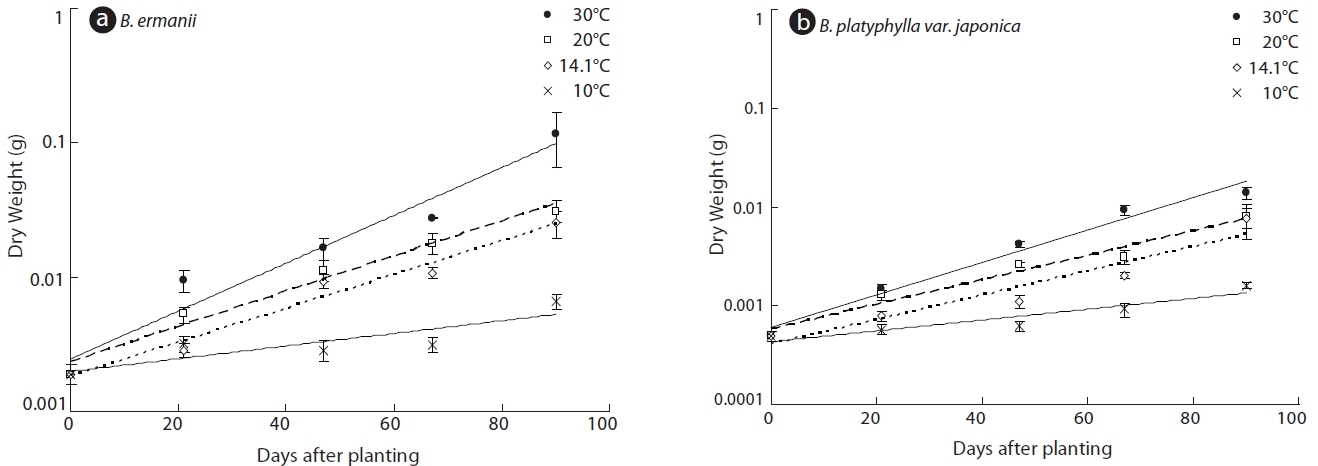
The dry weight of the B. ermanii and B. platyphylla var. japonica seedlings grown under different temperature conditions is given in Fig. 6; both species showed enhanced growth at higher temperatures. The relative growth rates (RGRs) at 14.1℃ (the mean annual temperature at 1,850 m during the growing season) were 0.031 g/day for B. platyphylla var. japonica and 0.029 g/day for B. ermanii, respectively, and these values did not differ significantly. The percentage survival of both species at constant temperatures of 10℃ and 14℃ and under natural conditions at different elevations on Mt. Neko are shown in Table 4a and 4b. The percentage survival of B. platyphylla var. japonica and B. ermanii seedlings was 50% and 100% at 10℃ after 90 days of planting. Conversely, at 14℃, percentage survival among seedlings was 80% in B. platyphylla var. japonica and 100% in B. ermanii (Table 4a). Under a constant temperature of 10℃, 75% of the B. ermanii seedlings were still healthy by 47 days after planting. Under natural condition, the survival rates of seedlings of B. platyphylla var. japonica were 10% at both 1,850 m and 2,050 m and of B. ermanii seedlings were 80% and 90% at the same elevations after 92 days of planting (Table 4b).
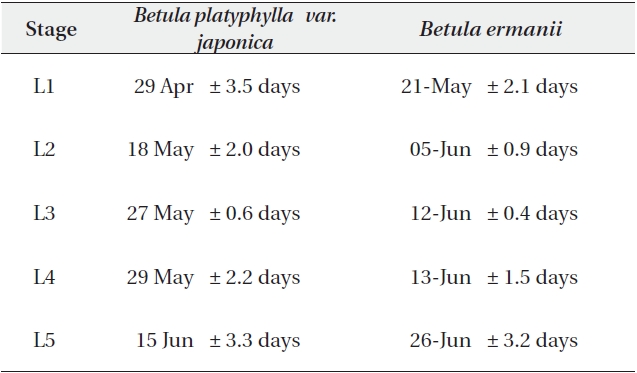
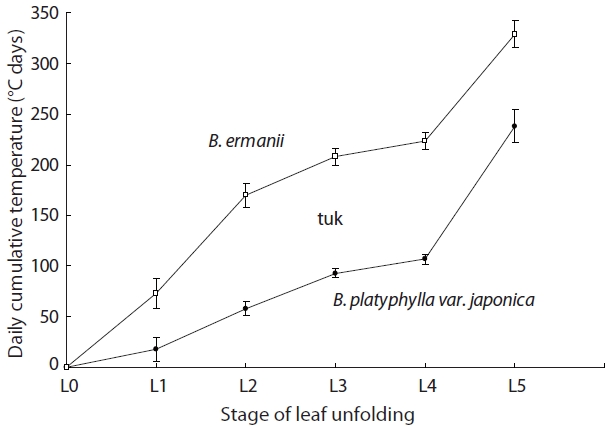
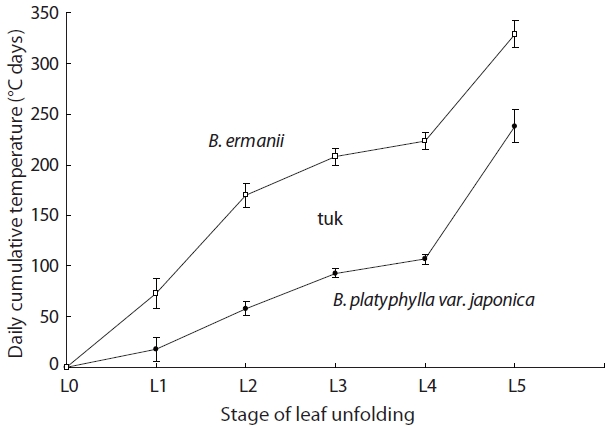
The stage of leaf unfolding in B. platyphylla var. japonica and B. ermanii, and the modified Kira's WI are shown in Fig. 7. Leaf expansion began (stage L2) at 58℃ day in B. platyphylla compared to 169℃ day in B. ermanii. B. platyphylla var. japonica required 240℃ day for full leaf expansion (stage L5) compared to 330℃ day for B. ermanii.
B. ermanii thus required higher daily cumulative temperatures for the initiation of leaf unfolding compared to B. platyphylla var. japonica, which meant that B. platyphylla var. japonica unfolded its leaves earlier than B. ermanii (Watanabe 1996). However, the WI of L4 stage needed 106℃ day in B. ermanii, whereas 131℃ day B. platyphylla var. japonica. Leaves of B. ermanii thus expanded comparatively quickly once unfolding started and attained full development. Compared to B. platyphylla var. japonica, it took 15 days longer for B. ermanii to pass from the L1 to the L2 stage, but 8 days less to pass from the L4 to the L5 stage (Kojima et al. 2003, Kato and Hayashi 2008).
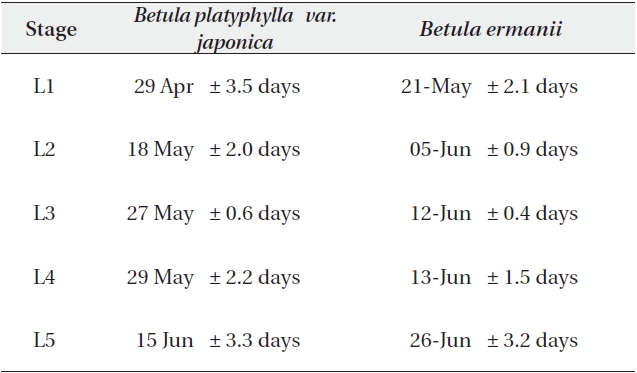
The dates when each leaf stage was reached at an altitude of 1,850 m are given in Table 5a. The L1 stage when the bud swells and the terminal end splits to expose the green tissues inside was reached on 29 April in B. platyphylla var. japonica and on 21 May in B. ermanii, a difference of more than 20 days. The L2 stage was reached on 18 May in B. platyphylla var. japonica and on 5 June in B. ermanii and the final stage (L5) was attained on 15 June in B. platyphylla var. japonica and on 26 June in B. ermanii.
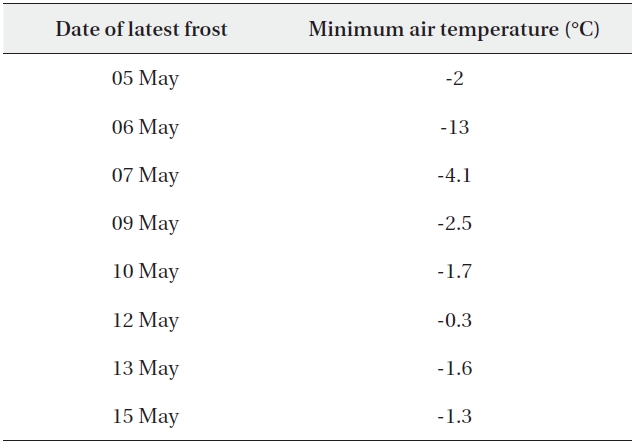
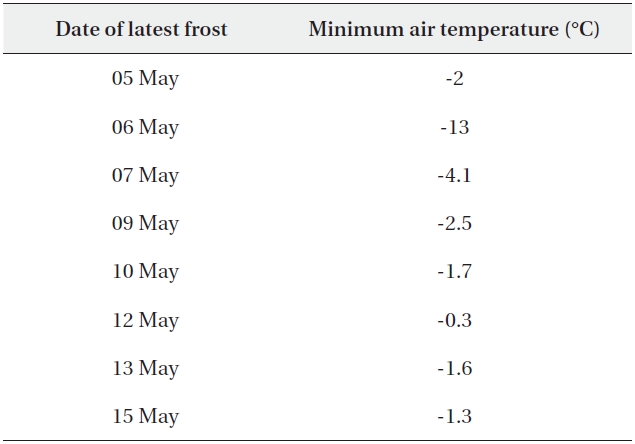
Table 5b shows the dates of late spring frosts at 1,850 m and the minimum air temperatures on those days. The last frost occurred on 15 May 1997, after the leaves of B. platyphylla var. japonica had begun to unfold. By this date B. platyphylla var. japonica was between the L1 and L2 stages, whereas B. ermanii was still in the winter bud stage (L0), implying that they escaped the potential damage associated with late spring frost.
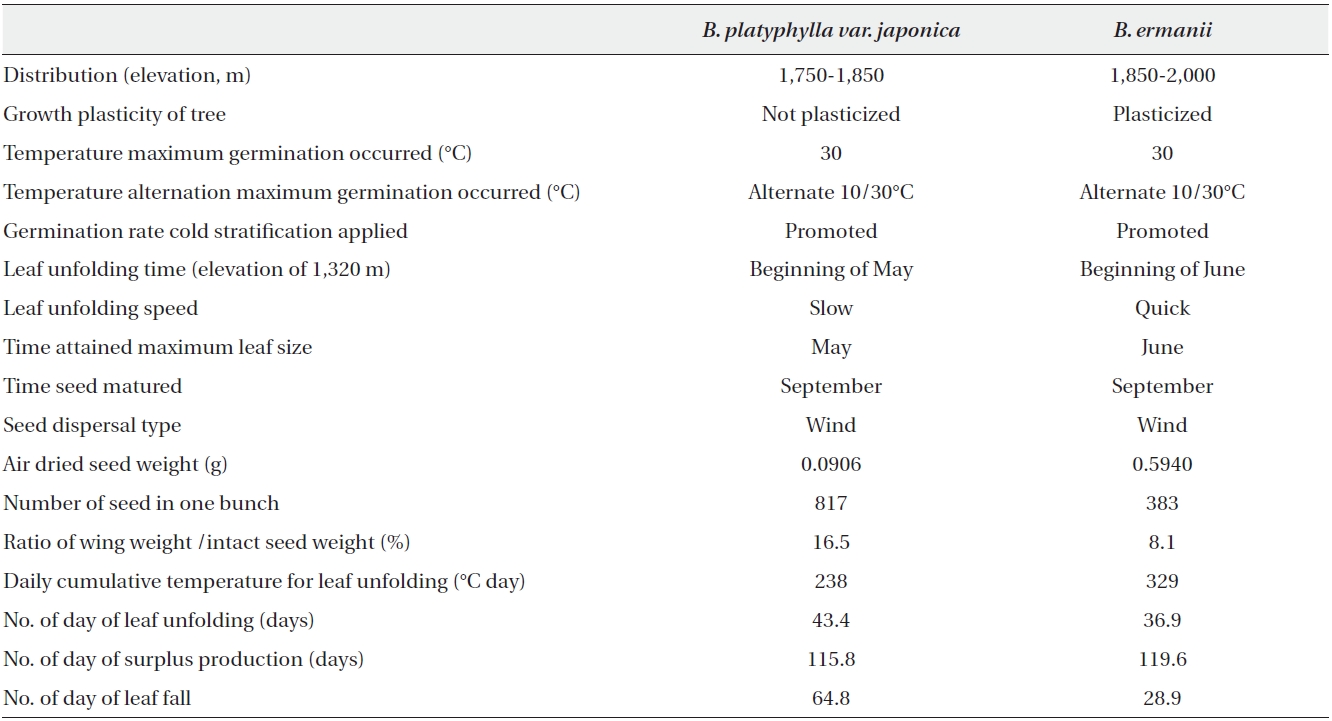
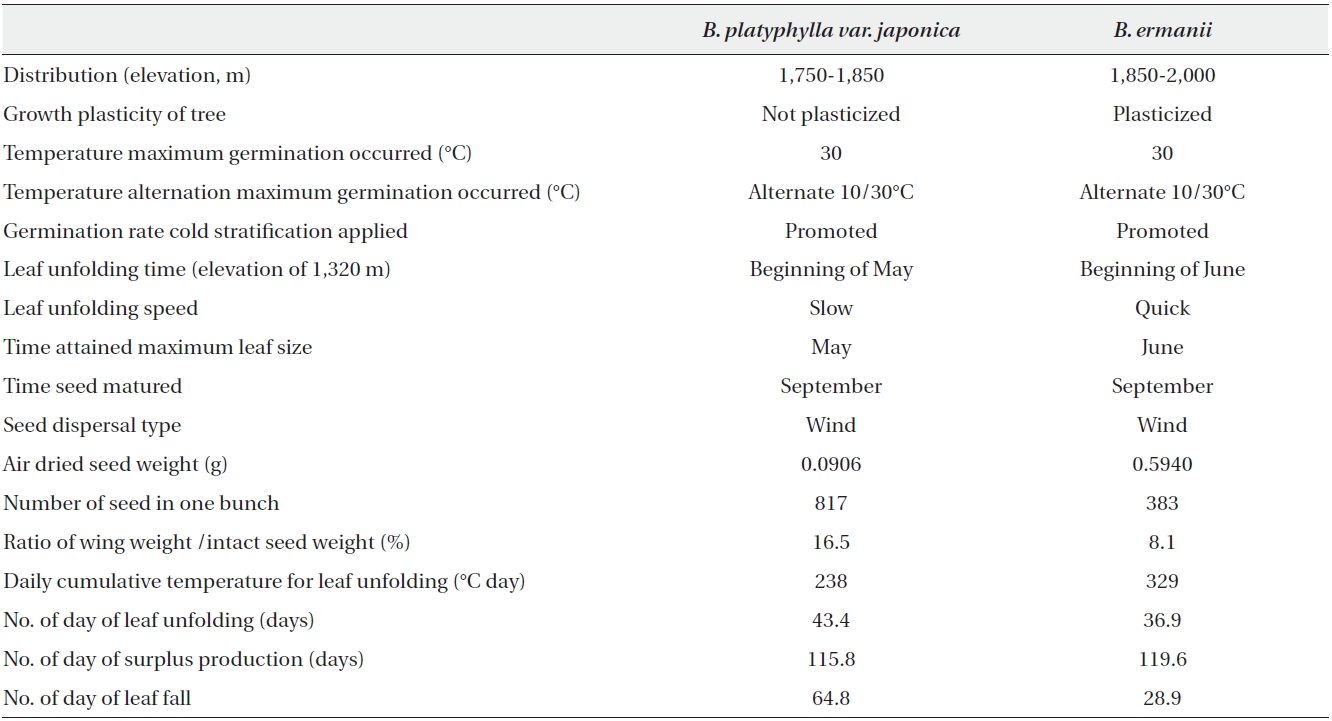
The biological characteristics of both species are summarised in Table 6. Briefly, B. ermanii germinates and/or grows at temperatures lower than those preferred by B. platyphylla var. japonica. Cold stratification for seeds and temperature alternation from 10 to 30℃ increased germination rates in both species. Non-stratified B. ermanii seeds did not germinate below 20℃, while germination occurred in B. platyphylla var. japonica at temperatures ranging from 10 to 30℃. The seedlings of both species grew best at 30℃. Below 14.1℃ the survival rate of B. ermanii was higher than that of B. platyphylla var. japonica under both experimental and field conditions. Leaf unfolding in B. platyphylla var. japonica commenced after 60℃ day of modified Kira's WI before the equivalent 160℃ day required for B. ermanii (Fig. 7). Finally, it was noted that the ratio of tree height versus DBH in B. ermanii varied considerably more than in B. platyphylla var. japonica with increasing elevation.
There are two possible hypotheses for the transition from B. platyphylla var. japonica to B. ermanii on Mt. Neko. The first hypothesis is that any discontinuous environmental conditions, including temperature in particular, exist along the altitudinal gradient. However, no such discontinuity was observed in either air or soil temperature gradients, or in mean air temperatures which decreased linearly with increasing elevation (Fig. 5). Thus, the first hypothesis was not supported by observed environmental conditions.
The second hypothesis was that these species respond differently to the same environmental cues due to their different biological characteristics such as timing of leaf unfolding, seed germination, frost avoidance and tolerance to heavy wind (Table 6) (Maruyama 1978, Linkosalo et al. 2000, Arora and Boer 2005). For example, leaf unfolding was initiated at a modified Kira's WI of 160℃ day in B. ermanii whereas leaf unfolding in B. platyphylla var. japonica occurred at 60℃ day at the same elevation. In other words, B. platyphylla var. japonica began leaf unfolding earlier than B. ermanii at the same elevation. At sites above 1,850 m, where late-spring frosts were recorded from 5 May until 14 May 1997 and WI was 60℃ day, the leaves of B platyphylla var. japonica were observed to have unfolded whereas the leaves in B. ermanii were still in winter buds. This meant that B. ermanii could avoid damage due to late frost while B. platyphylla var. japonica could not. No frosts occurred in the lower zone at this critical point in the growing season, which determines leaf unfolding in B. platyphylla var. japonica. Thus, the second hypothesis appears to offer a more likely reason for the transition between B. platyphylla var. japonica and B. ermanii at 1,850 m on Mt. Neko.
The principal difference in the vertical distribution of these two species was the timing of leaf unfolding and how this related to the occurrence of late spring frosts. According to field observations, frost damage to the young leaves of B. platyphylla var. japonica appeared to arrest leaf growth. However, during the same period, the leaves of B. ermanii were not damaged because the buds remained in their winter state until the risk of frost had passed.
Kojima et al. (2003) exposed buds of F. crenata in the winter bud stage (stage L0) and in early stage of leaf unfolding (stages L2-L3) to artificial frost. While the winter buds were not damaged by the frost and began unfolding normally afterwards, the L2-L3-stage buds did not unfold. This is likely to be the main reason why B. platyphylla var. japonica cannot become established above 1,850 m. On 5 May 1997, ice crystals were observed to have formed on the ends of upper branches of B. ermanii growing above 1,900 m. The frost did not damage the leaves of B. ermanii because they were protected by their winter buds, however, the unfolding leaves (stage L3) of B. platyphylla var. japonica were observed to be damaged.
The seeds of both species were produced in September (Table 6). Only a few seeds of B. ermanii germinate in autumn of the current year because they require moist stratification. These seeds contribute to the buried seed population and normally germinate in the following spring. On the other hand, the seeds of B. platyphylla var. japonica germinate soon after production, as they do not require stratification. The resulting seedlings cannot endure the cold winter conditions at high elevations and therefore cannot become established; in the areas where there is no late frost, such germination behaviour is considered advantageous. The seedlings of B. ermanii must be more tolerable for cold environment than that of B. platyphulla var. japonica because the seeds of former species are heavier than the latter species.
The thick, stunted growth form of B. ermanii trees growing near the summit of Mt. Neko, is well adapted to the strong winds and snow conditions (Okitsu and Satomi 1989). This morphological adaptation of trees to wind, which has been reported in several tree species (Larson 1965, Neel and Harris 1971, Lawton 1982), enables B. ermanii to survive on mountain summits (Ohsawa 1984) and is an important factor affecting the establishment of the B. ermanii zone in Hokkaido (Okitsu 1999). The absence of this form in B. platyphylla var. japonica further limits the success of this species in high mountain environments.
Taken together, these biological characteristics of B. platyphylla var. japonica and B. ermanii affect the transition between the two species at an elevation of 1,850 m on Mt. Neko, where the mean annual air temperature is 4℃. These same environmental factors, which favour the segregation of B. platyphylla japonica and B. ermanii at high elevations are also considered to affect the geographic distribution of these two species. Indeed, the 4℃ mean annual isotherm passes between Hokkaido and Sakhalin and coincides with the boundaries of their ranges (Kojima 1994). The distribution of B. ermanii on Hokkaido is primarily that area where the mean annual air temperature is 4.
On a large geographic scale, temperature is considered to be the primary environmental factor determining the vertical and horizontal distributions of these species. On a more local scale, the interrelationship between the biological and ecological characteristics of plants and their physical environments affect species segregation within a stand.