Seagrasses are a specialized group of flowering plants that are believed to have returned to the sea and adapted to living completely underwater (McComb et al. 1981, Robertson 1984). Seagrass meadows are among the most productive plant communities, and they provide habitats and shelter for commercially and ecologically valuable marine organisms (Holmquist et al. 1989, Montague and Ley 1993). Seagrass production significantly contributes to the abundance and production of a wide variety of organisms in coastal and estuarine ecosystems (Heck et al. 1995, Vizzini et al. 2002). Because seagrasses have high productivity, they require high nutrient inputs and play an important role in nutrient cycling in these ecosystems (Blackburn et al. 1994, Park et al. 2009). Thus, accurate assessment of seagrass productivity is critical to understanding the ecological role and value of seagrass beds.
Numerous methods have been developed to measure seagrass productivity (for a review, see Kemp et al. 1990). The leaf marking method developed by Zieman (1974, 1975) has been widely used to assess leaf productivity and was improved by several researchers (Sand-Jensen 1975, Kentula and McIntire 1986, Roman and Able 1988, Dennison 1990a, Ibarra-Obando and Boudouresque 1994). However, this method can produce underestimatesdue to its failure to consider new leaf growth within the sheath below the reference mark (Sand-Jensen 1975, Ibarra-Obando and Boudouresque 1994), damage to the leaf tissues caused by marking (Dennison 1990a), and its disregard for the leaf maturation process (Bedhomme et al. 1983. Short 1987, Gaeckle and Short 2002). Thus, a new productivity assessment method, the elongationmass method (Short ’87 method), in which both leaf elongation and weight gain are considered, has been developed to reduce the downward bias in seagrass leaf productivity resulting from weighing immature leaf sections from newly grown leaf tissues (Short 1987, Gaeckle and Short 2002). In this method, leaf productivity is calculated by multiplying the leaf elongation rate by the leaf weight-to-length ratio (mg cm?1) of mature leaf material to account for the leaf weight gain that is part of the total leaf growth; thus, this method can reflect leaf maturation processes (Gaeckle and Short 2002).
The plastochrone method was developed to complement previous ways of measuring seagrass productivity (Short and Duarte 2001). The leaf plastochrone interval is defined as the period between successive initiations of two leaves on a shoot (Patriquin 1973, Sand-Jensen 1975, Jacobs 1979). In the plastochrone method, leaf productivity is calculated based on the plastochrone interval and the weight of a mature leaf. This method was described as a simple and accurate technique for assessment of seagrass productivity (Short and Duarte 2001, Gaeckle and Short 2002, Lee 2004). Although seagrass productivity has been measured using these methods worldwide, few studies have compared these three measuring methods. In the present study, we compared three measurement methods, the commonly used leaf marking method, the elongation-mass method, and the plastochrone method, to determine which is the more accurate and reliable technique for measuring seagrass productivity.
Our study was conducted in Koje Bay (N 34o48?, E 128o35?) on the southern coast of the Korean peninsula (Fig. 1). Four
>
Leaf marking and leaf productivity estimates
We estimated leaf elongation, leaf plastochrone interval and leaf productivity in each season (October 2004 and January, March and June 2005). Thirty to forty shoots of
For the conventional leaf marking method, leaf material was separated into leaf tissue produced before and after marking. Any epiphytes and sediments were removed from these leaves by gentle scraping. Separated leaf materials were dried at 60℃ to a constant weight. Leaf productivity (mg dry weight shoot-1 day-1) was estimated using the conventional leaf marking method by dividing the dry weight of new leaf tissue produced after marking by the number of days since marking.
For the elongation-mass method, the total length of new leaf tissues produced after marking was measured. Additionally, a weight-to-length ratio (mg cm-1) was measured using 20-30 cm of mature leaf material (generally the third leaf). The mature leaf material was cleaned of epiphytes and dried at 60℃ to a constant weight; then the weight-to-length ratio was calculated. Leaf productivity was estimated using the elongationmass method by multiplying the weight-to-length ratio by the length of new leaf material produced after marking. For the plastochrone
For the plastochrone method, the leaf plastochrone interval was calculated by dividing the marking period by the number of new leaves produced after marking. The dry weights of the youngest mature leaf and the rhizome/ root segments from the first to sixth youngest nodes were measured at every sampling interval. The youngest mature leaf was generally the third leaf and the youngest mature rhizome segment was usually the longest one. Leaf and below-ground productivities of each shoot were calculated using the following equations:
Leaf productivity (mg dry weight shoot-1 day-1) = dry weight of the youngest mature leaf (mg dry weight shoot-1) / the plastochrone interval (days) Below-ground productivity (mg dry weight shoot-1 day-1) = dry weight of a mature rhizome/root segment (mg dry weight shoot-1) / the plastochrone interval (days)
All data are presented as means ± SE. Data were tested for homogeneity and normality of variances to meet the assumptions of parametric statistics. If these assumptions were violated, data were log-transformed. We assessed the significance of the differences in leaf elongation and leaf plastochrone interval among seasons using one-way ANOVA. We also tested for significant differences in leaf productivity estimates using the three measurement methods in each season and compared total, aboveground and below-ground productivities among seasons with one-way ANOVA. When a significant difference among the variables was observed, the means were compared using the Student-Newman-Keuls test to determine which of the treatments differed significantly. We also compared the slopes of the linear regressions between leaf productivity estimates using the three measurement methods. An alpha level of 0.05 was used for statistical tests. All data analyses were performed using SPSS (version 12.0).
>
Leaf elongation and leaf plastochrone interval
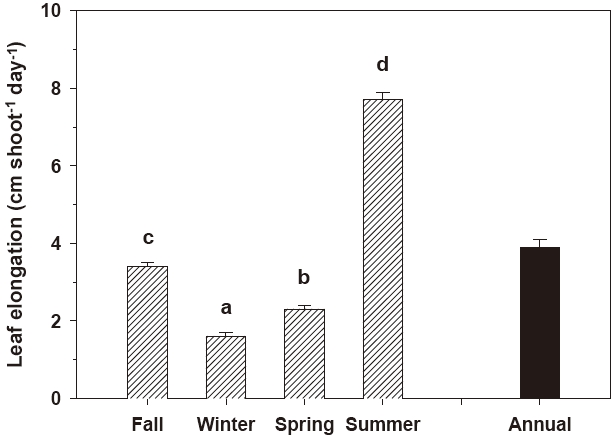
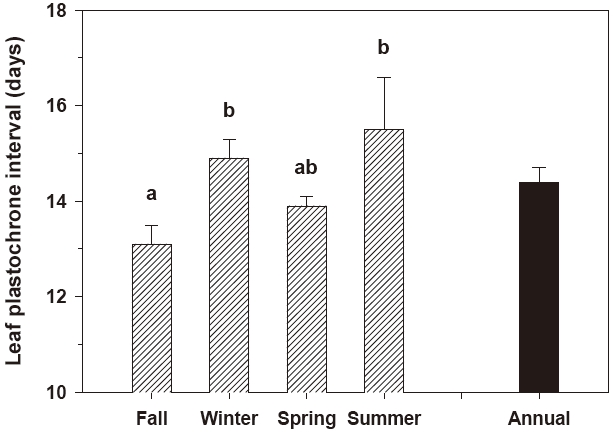
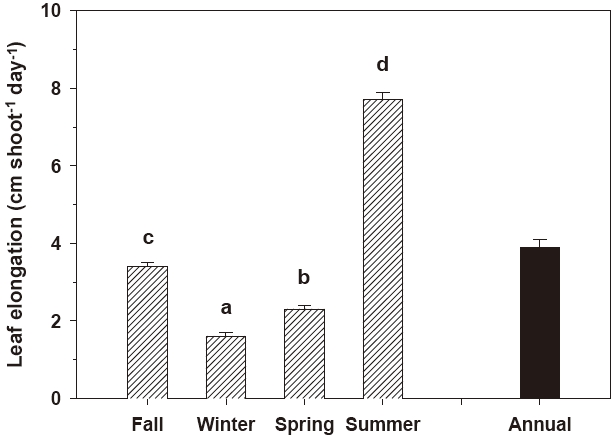
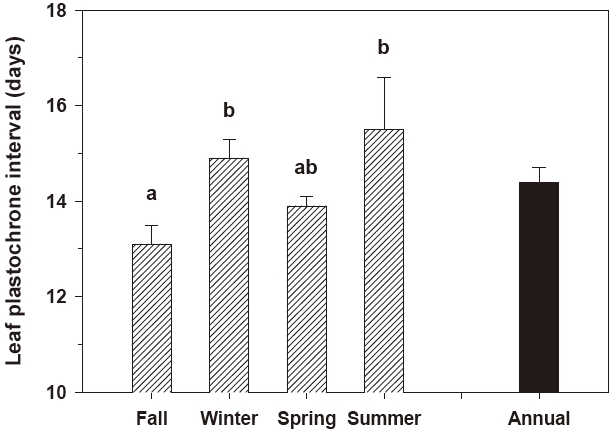
Leaf elongation exhibited significant (P < 0.001) seasonal variation (Fig. 2). The rate of leaf elongation ranged from 1.6 cm shoot-1 day-1 in January 2005 to 7.8 cm shoot-1 day-1 in June 2005; the annual mean was 3.9 ± 0.2 cm shoot-1 day-1. The leaf plastochrone interval also varied significantly (P = 0.045) among seasons. The leaf plastochrone interval was highest (15.5 ± 1.6 days) in summer and lowest (13.1 ± 0.4 days) in fall (Fig. 3). The annual mean leaf plastochrone interval was 14.4 days.
>
Comparisons of the three measurement methods
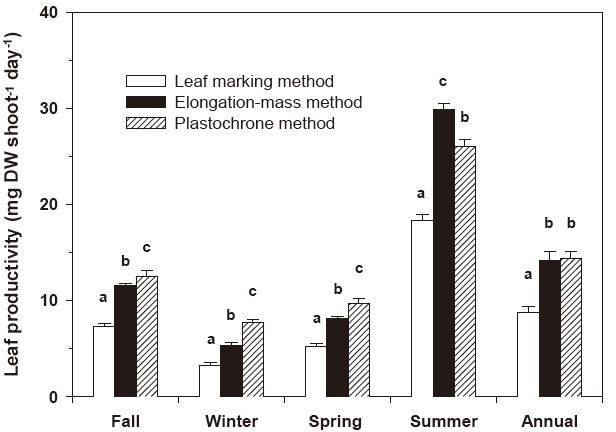
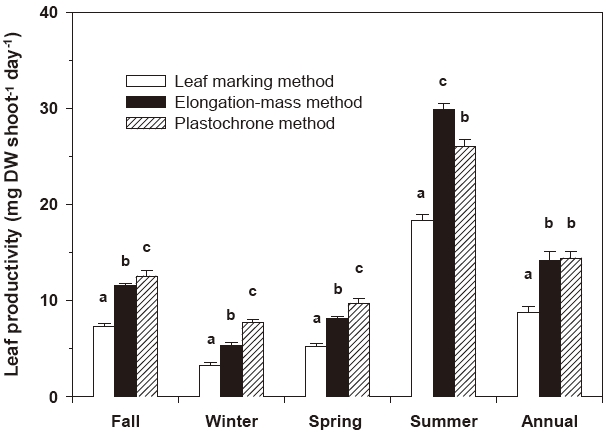
Leaf productivity estimates produced with the three measurement methods (the conventional leaf marking method, the elongation-mass method and the plastochrone method) differed significantly throughout the experimental periods (Fig. 4). Leaf productivities estimated using the plastochrone method were significantly higher (P < 0.001) than those estimated using the conventional leaf marking method and the elongation-mass method in all experimental seasons except summer (Fig. 4). During fall, winter, and spring, the values derived using the plastochrone method were 1.7-2.4 times higher than those produced using the conventional leaf marking method and 1.1-1.6 times higher than those derived using the elongation-mass method.

Above-ground, below-ground, and total shoot productivities (mg dry weight shoot-1 d-1) of Zostera marina estimated using the plastochrone method.
Leaf productivity estimates produced using the elongation-mass method were significantly higher (P < 0.001) than those estimated using the conventional leaf marking method throughout the experimental periods and were also higher than the plastochrone-methodbased estimates in summer, when leaf elongation was highest (Fig. 4). The conventional leaf marking method consistently produced the lowest leaf productivity estimates. On an annual basis, leaf productivity estimates produced with the conventional leaf marking method were 38% and 39% lower than those produdced with the elongation-mass method and the plastochrone method, respectively (Fig. 4).
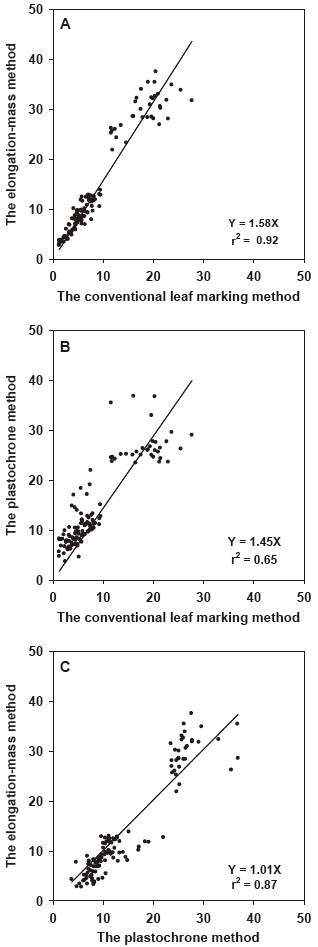
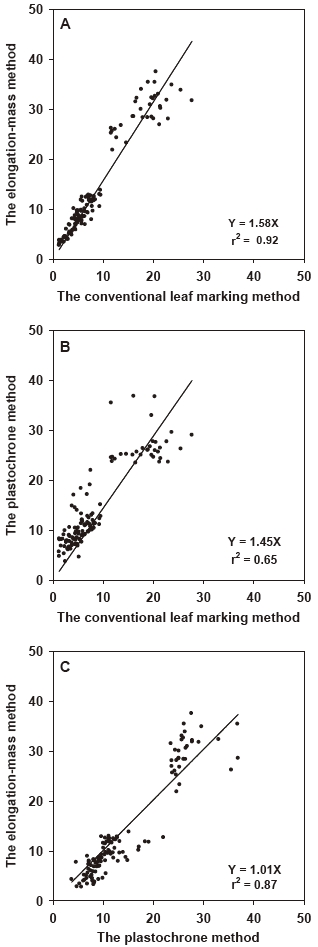
Estimates of leaf productivity using the three measurement methods were strongly correlated (Fig. 5). Estimates produced using the conventional leaf marking method exhibited the strongest relationship with those from the elongation-mass method (r2 = 0.92) and a weaker relationship with those using the plastochrone method (r2 = 0.65). Estimates produced using the elongation-mass method also showed a high and significant correlation (r2 = 0.87) with those produced using the plastochrone method.
>
Below-ground productivity using the plastochrone method
Below-ground productivity was relatively constant during all seasons except summer (Table 1). Belowground productivity was highest in summer (5.73 mg dry weight shoot-1 day-1) and lowest in winter (3.29 mg dry weight shoot-1 day-1). Below-ground production accounted for 24.4% of total production on an annual basis. Total (above- + below-ground) productivity varied significantly (p < 0.001) with season (Table 1). Total productivity was highest in summer (31.76 mg dry weight shoot-1 day-1) and lowest in winter (10.96 mg dry weight shoot-1 day-1).
The leaf marking technique described by Zieman (1974, 1975) has been the predominant method used to assess the leaf productivity of seagrasses over the past 30 years. However, several researchers have pointed out inherent problems with the conventional leaf marking method (Sand-Jensen 1975, Brouns 1985, Ibarra-Obando and Boudouresque 1994, Gaeckle and Short 2002). The leaf marking method ignores maturation of leaf tissues located above the marking point (Bedhomme et al. 1983, Short 1987, Gaeckle and Short 2002), and new leaf growth occurring below the marking point may also be excluded in the conventional leaf marking method (Sand-Jensen 1975, Brouns 1985, Ibarra-Obando and Boudouresque 1994). In this study, the conventional leaf marking method resulted in the lowest leaf productivity values among the three measurement techniques. On an annual basis, leaf productivities estimated by the conventional leaf marking method were only 62% and 61% of those estimated using the elongation-mass and the plastochrone methods, respectively. Regressions among estimates produced using the three measurement methods also showed that the conventional leaf marking method produced the lowest estimates of leaf productivity. Thus, as many researchers have pointed out, productivity estimates produced using the conventional leaf marking method probably underestimate actual productivity due to the failure to include leaf maturation processes and new leaf growth within the sheath.
Since the elongation-mass method incorporates leaf maturation processes, this technique is considered to be a more complete productivity measurement modality than the conventional leaf marking method (Gaeckle and Short 2002). Although the elongation-mass method can reflect leaf tissue maturation processes, this method still ignores new leaf growth below the marking points. On an annual basis, leaf productivity estimates using the elongation-mass method were approximately 98% of those attained with the plastochrone method. However, the elongation-mass method estimated 15% higher leaf productivity than the plastochrone method during summer, when leaf elongation rates increased dramatically.
Leaf productivity usually exhibits clear seasonal variation, increasing during spring and early summer, and decreasing during fall and winter (Lee et al. 2005). During summer, leaf maturation processes appeared to be at a minimum as leaves elongated greatly (Park 2008). Since completely mature leaf tissues were used to calculate leaf productivity with the elongation-mass method, the productivity estimate produced using this technique may be an overestimate during seasons of maximum leaf elongation. Gaeckle and Short (2002) also reported that productivity estimations generated using the elongationmass method were higher than those made using the plastochrone method in July.
Of the three measurement methods, the plastochrone method generated the highest leaf productivity estimates on an annual basis. The leaf productivity estimates made with the plastochrone method were 64% and 2% higher than those obtained with the conventional leaf marking method and the elongation-mass method, respectively. Leaf productivities estimated using the plastochrone method reflect both leaf maturation and growth within the sheath. Assessment of belowground productivity is remarkably difficult (Dennison 1990b). Rhizome tagging has been used to estimate below-ground productivity (Dennison et al. 1985), but tagging rhizomes requires substantial effort under field conditions. Below-ground production has been assessed as 15-43% of the total seagrass production (Nienhuis and de Bree 1980, Robertson and Mann 1984, Park et al. 2009). We estimated below-ground productivities using the plastochrone method as 3.29-5.73 mg dry weight shoot-1 day-1, which accounted for about 18-30% (mean = 24%) of total eelgrass productivity. In Koje Bay, leaf biomass ranged from 12 g dry weight m-2 in December to 696 g dry weight m-2 in June, whereas below-ground biomass ranged from 7 g dry weight m-2 in March to 92 g dry weight m-2 in August (Lee et al. 2005). Below-ground biomass accounted for about 20% of the total biomass. Thus, below-ground productivity in this study was consistent with the pattern of biomass allocation. These high contributions of below-ground productivity to total seagrass production imply that below-ground production should not be ignored when attempting to clarify seagrass production dynamics. Below-ground productivity as well as above-ground productivity can be easily estimated using the plastochrone method. Thus, the plastochrone method is an effective assessment modality for total seagrass productivity.
In conclusion, the results of our comparisons of the three measurement methods suggest that the conventional leaf marking method underestimates eelgrass leaf productivity as it ignores leaf maturation processes and new leaf growth within the sheath. Since the elongation-mass method can reflect leaf maturation processes, it appeared to provide a more reliable leaf productivity estimate than the conventional leaf marking method. However, the plastochrone method appears to be the most effective assessment technique for measuring both above- and below-ground productivities.