Intertidal regions have been considered as highly productive systems in the marine environments (Valiela and Teal 1979). In the intertidal flats, the benthic microalgae play an important role as primary producers (Sullivan and Daiber 1975; Rizzo and Wetzel 1985; Underwood and Kromkamp 1999). This primary production of organic compounds is the main food sources for microbial, microbenthic, meiobenthic and macrobenthic organisms abundant in the intertidal flats (Heip et al. 1995; Kang et al. 2003). The primary production contributes to a substantial proportion of the energy flow in estuarine systems (Herman et al. 1999). In addition to supplying a food resource, microalgae excrete high molecular weight polysaccharides that contribute to sediment stabilization and reduce sediment resuspension during both tidal emergence and storms (Lucas et al. 2003). This protective layer also prevents underlying heterotrophic bacteria from being removed by abrasion (Miller et al. 1996).
In nutrient dynamics of the intertidal ecosystems, sediments play a role as source through regeneration to the water column deposited on the bottom (Hopkinson 1987) and as sink through benthic nitrogen retention processes including burial in deeper sediment layers and denitrification (Smith et al. 1985). Benthic algae can significantly reduce the release of nutrients from the sediments by utilizing the nutrients when they diffuse across the surface and indirectly by oxygenating surface sediment (Andersen et al. 1984; Sundback and Graneli 1988; Rizzo et al. 1992; Tyler et al. 2003; Sundback et al. 2004; Sin et al. 2006).
The intertidal flats are located mainly in the west coasts of Korea (83%) and especially 41% of the intertidal flats located in the west and south coasts of Jeonnam Province (http://www.wetland.or.kr). Various studies have been reported focusing on dynamics of sedimentation (Choi 1992; Ryu et al. 2003; Ryu and Sin 2006), physical- chemical properties (Kim and You 2001; You et al.2003), macro-sized benthos (Koh and Shin 1988; An and Koh 1992; Lim et al. 1997) for intertidal flats in Korea. Variations of benthic microalgae biomass and community were investigated for Kyonggi Bay (Yoo and Choi 2005), Kuem River (Kim and Cho 1985), Saemangeum (Oh and Koh 1995; Oh et al. 2004), Youngkwang and Gangjin (Lee 2002; Lee 2003). However, dynamics of benthic microalgae were not explored for the intertidal flats of Kwangyang Bay located in the south coast.
The loss of intertidal flats in Kwangyang Bay has increased since 1970s due to reclamation of shallowwater area for construction of large-scaled port and industry facilities. Water surface area was 233 km2 in 1973 and decreased to 193 km2 in 1990 (17% loss) and to 165 km2 (30% loss) in 2003 (Ryu 2003; Choi et al. 2003). The intertidal flats are currently remained in the northern and southern areas of Song Island. Although the results have been reported on trophic importance of benthic microalgae in the marine food web of the Kwangyang Bay using isotope analyses (Kang et al. 2001; Kang et al. 2003), variations of benthic microalgal biomass and controlling mechanisms related to the variations were not documented. The goal of this study was to investigate temporal and spatial distributions of benthic microalgae and physical-chemical properties of sediments for better understanding dynamics of benthic microalgae in the intertidal flats of Kwangyang Bay.
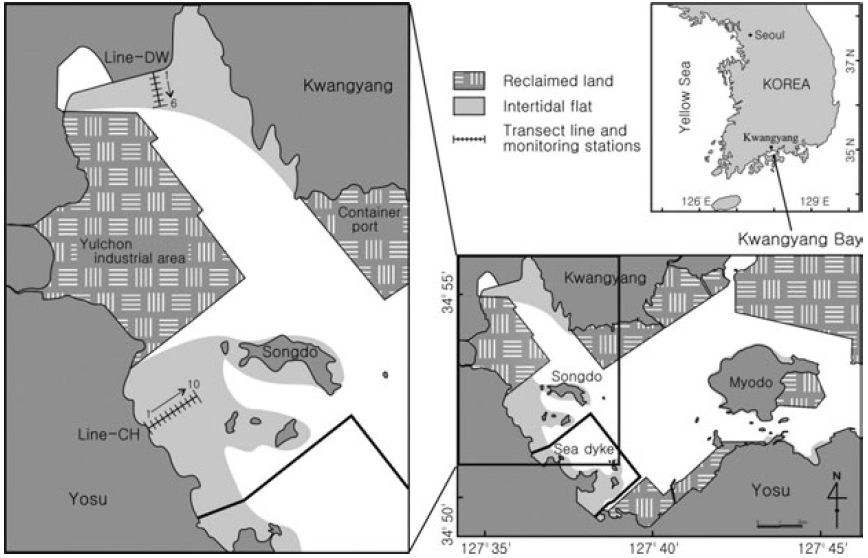
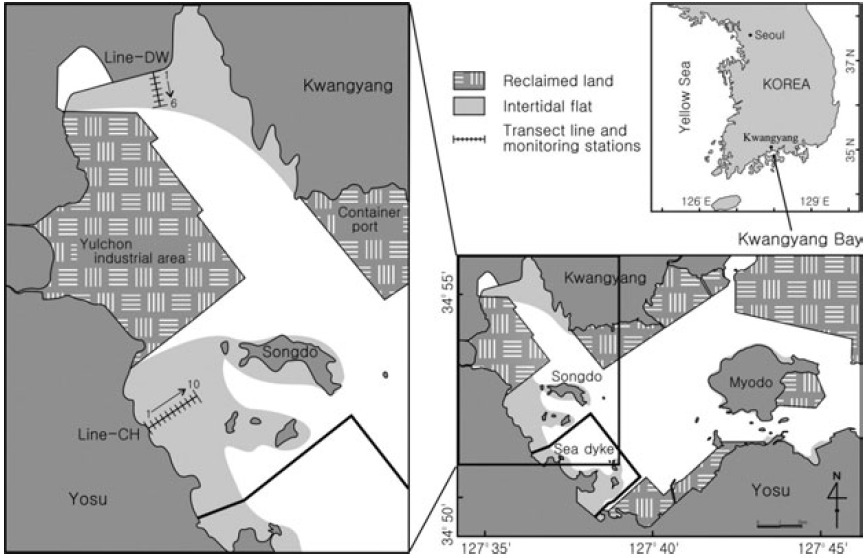
The Kwangyang Bay is located in the central region of South Coast and a semi-closed system surrounded by Yosu Peninsula and Namhae Island (Fig. 1). Study sites were selected from the intertidal flats in the range of 127° 34’-127° 39’ E, 34° 51’-34° 56’ N. The Kwangyang port (container) and Yulchon industry clusters are located in the east and west of the study sites and a sea dyke was constructed for reclamation of coastal regions for 2 years from 2003 to 2004 (Fig. 1). The intertidal flats in west of Kwangyang Bay were mainly reclaimed and study sites are the flats remained in the Kwangyang Bay. Dongcheon stream is extended in the north of study sites and freshwater is introduced especially during wet season. Main tidal channel is located between the Kwangyang Port and Yulchon industry clusters and has been dredged to maintain c.a. 7 m depth for transportation of vessels in the area.
Sediment types of the study sites are grouped in three classes including muddy sand, sandy mud and mud (Ryu 2003). Sediments in the northern regions of intertidal flats (Line-DW) are mainly composed of muddy sand whereas sediments in the western regions (Line- CH) are primarily sandy mud. Muds are abundant in the subtidal zones. Annual average of air temperature and rainfall of the regions is 14.1°C and 1893.5 mm respectively for 10 years from 1991 to 2000. The hydrological data presented in the study were collected by Korea Meteorological Administration.
Samples were collected at the beginning of low tide when the flats were exposed to air in June, July, September, November in 2004 and February and May in 2005. Stations were selected at interval of 100 m from upper to lower regions of the intertidal flats in Dowal-Ri (Line-DW) and Chohwa-Ri (Line-CH) (Fig. 1). Seasonal characteristics were investigated by classifying June-July as summer and September-November as fall. February and May were classified as winter and spring respectively.
Temperature of sediments was measured after inserting the mercury thermometer to the depth of 5 cm in the sediments for 1 or 2 minutes. Interstitial water of surface sediments (1 cm) was collected in-situ to measure ambient nutrients concentrations using a 100 ml syringe. The interstitial water was filtered using 25 mm Acrodisc Syringe Filters (0.45 μm) and inserted into HDPE bottle for freezing. Ambient concentrations of nutrients including nitrite+nitrate (NO2 ? + NO3 ?), ammonium (NH4 +), orthophosphate (PO4 ?3), and dissolved silica (DSi) were measured by the nutrient autoanalyzer (Bran Luebbe). Sediment cores (replicates) were collected using polycarbonate core tube (5 cm x 10 cm) to measure properties of sediments such as water contents (%) and density (mg cm?3) and chlorophyll a in the surface sediments. Sediment samples were extruded gravimetrically and sectioned at various depth using a sharp knife. Sediment bulk density for aerobic (surface) and anaerobic (subsurface) layers of two cores was determined based on volume (ml) and dry weight (g) of sediments in the cores. Aerobic and anaerobic layers were identified by colors of layers. The sediments were dried at 50°C for 5-7 days to obtain the dry weight. The porosity of the segments was the mass difference between wet and dry sediments. One segment (0-0.5 cm depth) of the two cores was analyzed for chlorophyll a (Arar and Collins 1992). Sediment chlorophyll a was measured by taking replicate samples from each segment using 2 cm diameter core tubes to a depth of at least 0.5 cm. Samples were extracted with 5 ml acetone (90%) in the dark for 24 hours analyzed by a Turner Designs Model 10 Fluorometer (Arar and Collins 1992). Surface sediments were collected using a plastic scoop and analyzed grain size by sieve procedure (Ingram 1971). The ratios of fluorescence before and after acidification (chlorophyll: pheopigment) were analyzed as a proxy for grazing effect (top-down control) since herbivores convert chlorophyll a to pheopigments, which are released as egested fecal material. The ratio of chlorophyll a and pheopigments, calculated from the ratios of fluorescence before and after acidification is an indirect measure of grazing activity (e.g. Welschmeyer and Lorenzen 1985); the lower the ratio, the higher the grazing activity.
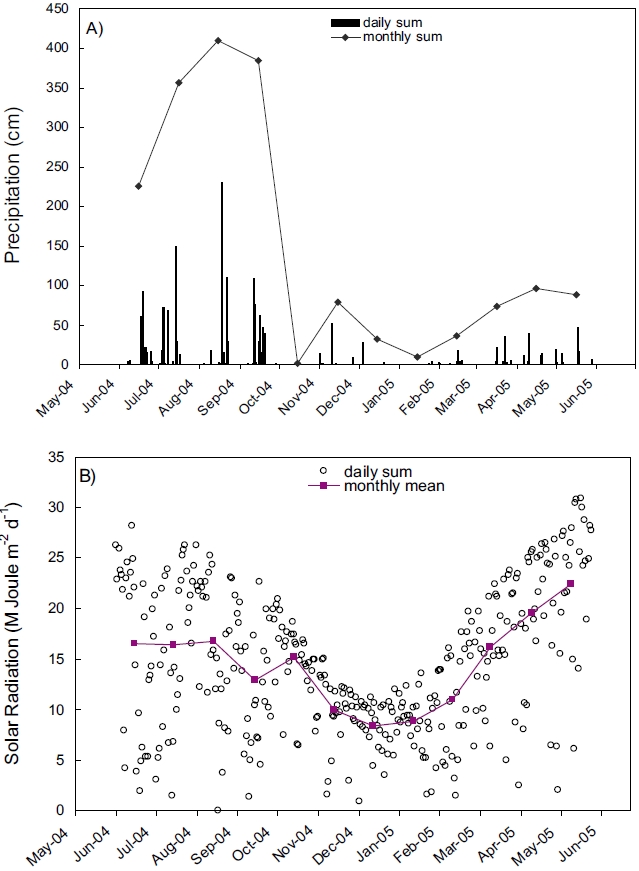
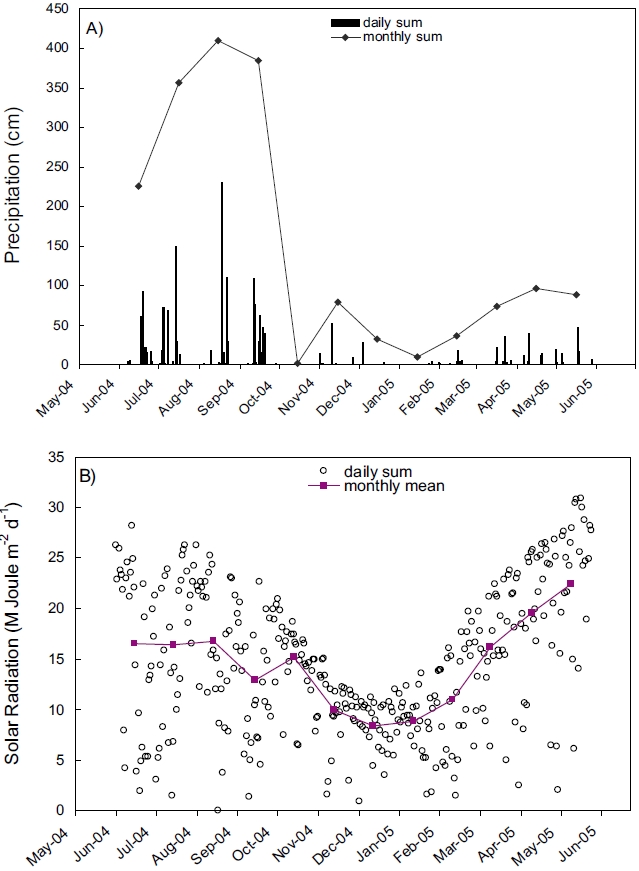
Records of rainfall and solar radiation collected near the Kwangyang Bay area were presented in Figure 2. Precipitation was extraordinarily high during summer compared with other seasons indicating that the study area is influenced by Monsoon climate (Fig. 2A). Solar radiation was also high during summer and low during winter similar to the annual patterns of other areas in mid-latitude (Fig. 2B).
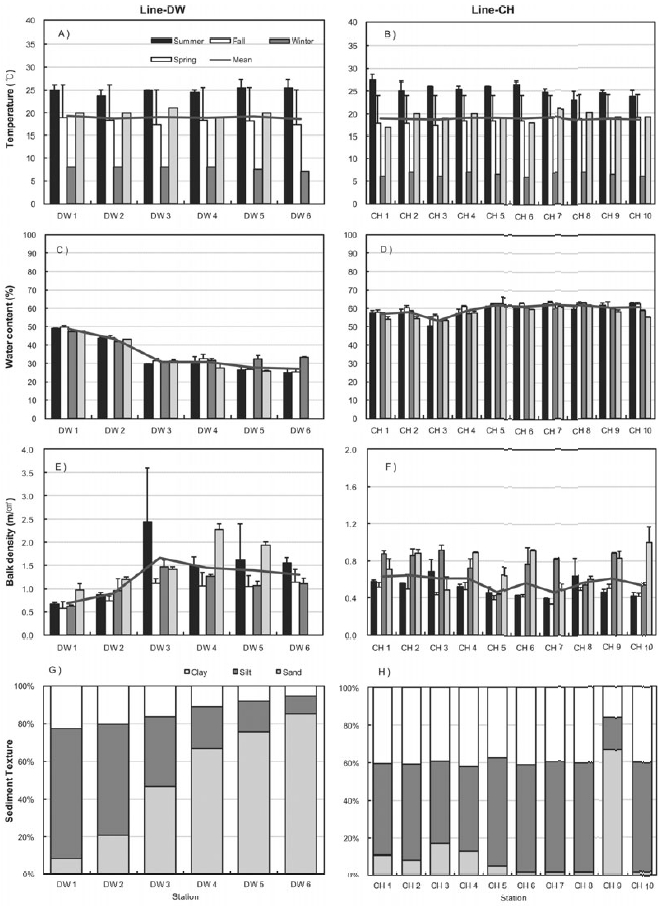
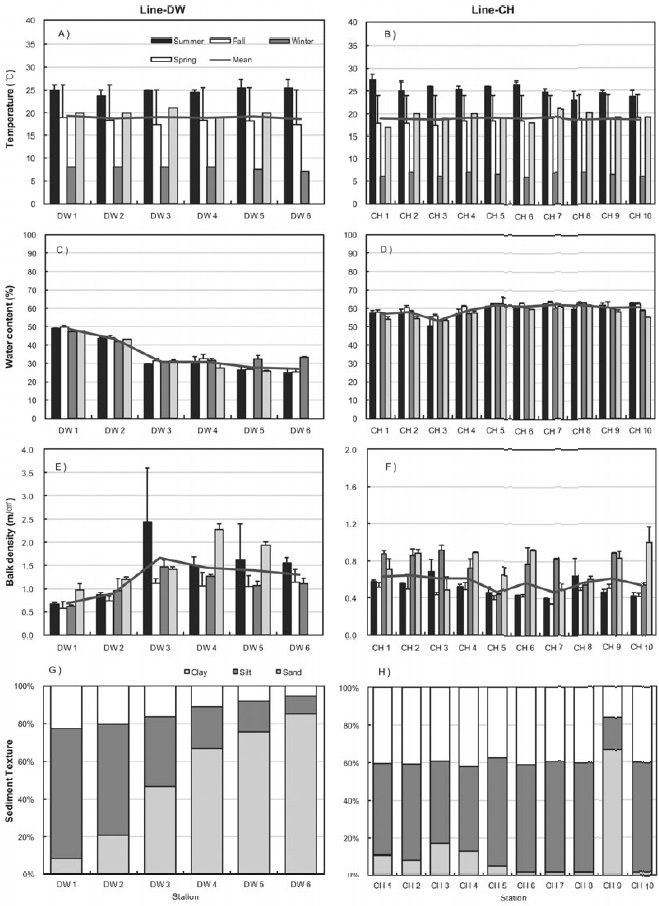
Figure 3 shows the seasonal characteristics of sediments including sediment temperature, water contents, bulk density and sediment texture at the sampling sites of Line-DW and Line-CH. Temperature was high in summer and low in winter and spatial difference was not high between the sampling sites (Figs 3A and 3B). Sediment temperature in the Line-CH was slightly higher than in the Line-DW. Seasonal variation of water contents in the study sites was not evident but water contents decreased as we approach seaward in the Line-DW whereas water contents were remained similar as seaward in the Line-CH (Figs 3C and 3D). Sediment bulk density was generally high during spring and summer whereas density was low during fall and winter in Line- DW (Fig. 3E). In Line-CH bulk density of sediments was relatively high during winter and spring and low during summer and fall (Fig. 3F). Bulk density was mostly low at the landward sites compared with that of seaward sites in Line-DW whereas spatial variation was not evident in Line-CW. Silt and clay were dominant in the sediment texture of the sites near lands but sediment texture was shifted to sand-dominant texture (coarse) as seaward in Line-DW (Fig. 3G). Silt and clay (fine) were dominant in sediment texture of Line-CH except Station CH9 where sand was dominated (Fig. 3H).
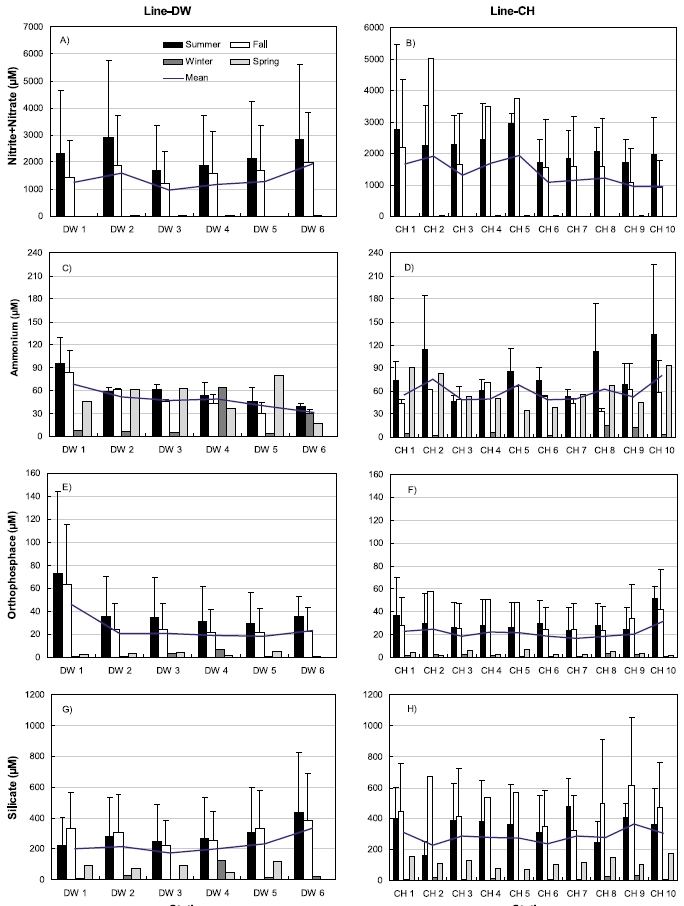
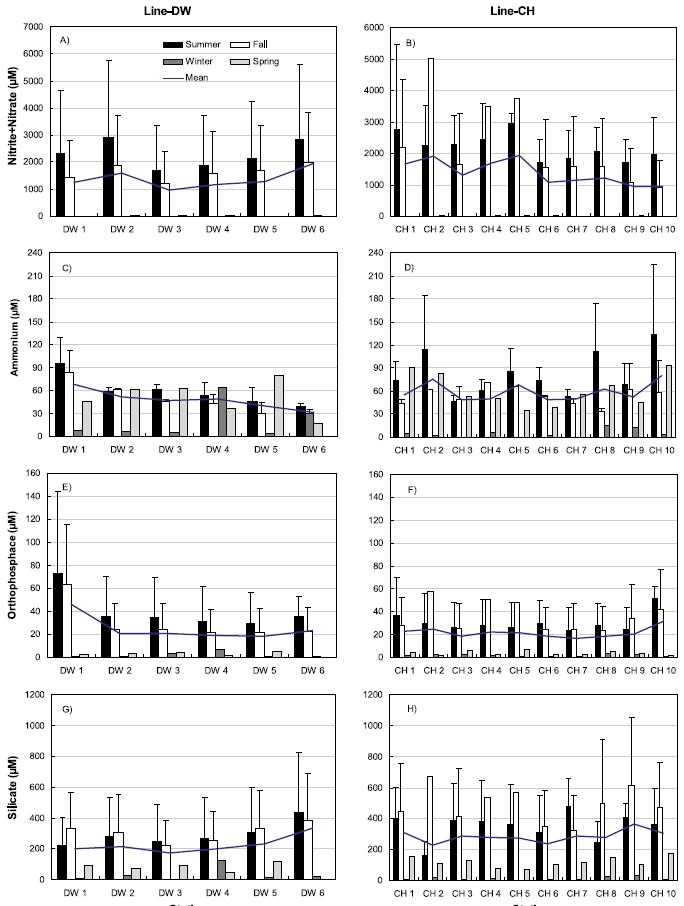
Nutrient concentrations in the sediments including nitrite+nitrate, ammonium, orthophosphate, silicate were shown in Fig. 4. Nitrite + nitrate concentrations were highest during summer and lowest during winter ranging from 1.67 μM (February) to 5767.2 μM (July) in Line-DW (Fig. 4A). Similar pattern was observed for nitrite+nitrate ranging from 0.57 μM (February) to 5461.2 μM (July) in Line-CH (Fig. 4B). No evident spatial variation was observed in Line-DW but the concentration slightly decreased as seaward in Line-CH. Ammonium concentrations were high during spring, summer and fall whereas the concentrations were low during winter in the Line-DW except Station DW4 (Fig. 4C) and similar seasonal pattern was observed in the Line-CH (Fig. 4D). Ammonium concentrations were decreased as seaward in Line-DW whereas no clear trend was detected in Line- CH. Orthophosphate concentrations were highest during summer and lowest during winter ranging from 0.68 μM (February) to 143.9 μM (July) in Line-DW except DW4 (Fig. 4E). Similar pattern was observed for orthophosphate ranging from 0.49 μM (February) to 76.4 μM (September) in Line-CH (Fig. 4F). The concentration in Line-DW was slightly decreased as seaward but no evident spatial variation was observed for Line-CH. Silicate concentrations were highest during summer or fall and lowest during winter except Station DW4 ranging from 3.4 μM (February) to 830.7 μM (July) in Line-DW (Fig. 4G). Silicate was maximum mostly during fall and mini-mum during winter ranging from 3.3 μM (February) to 1052.2 μM (June) in Line-CH (Fig. 4F). No evident spatial variations were observed in Line-DW and Line-CH.
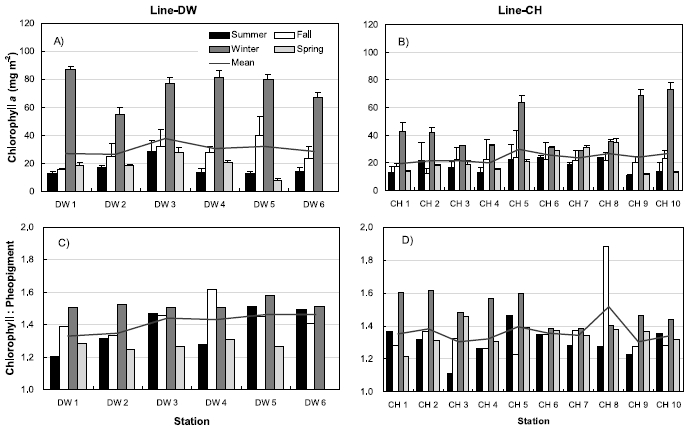
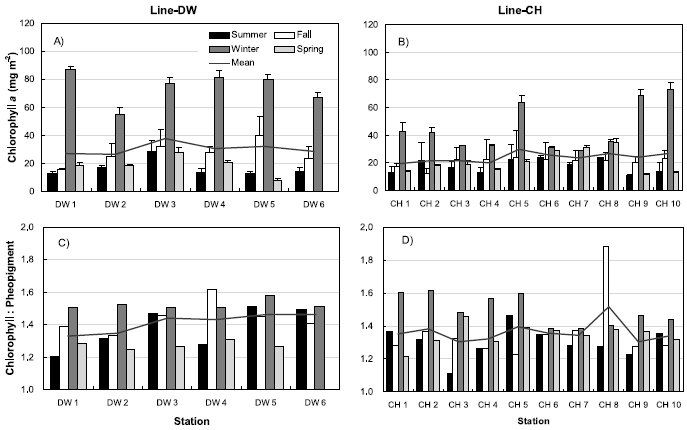
Figure 5 shows the seasonal variations of sediment chlorophyll a and the ratios of chlorophyll:pheopigment from the sampling sites in the Line-DW and Line-CH. Sediment chlorophyll a concentrations were maximum during winter and minimum during summer or spring (Fig. 5A) ranging from 8.3 mg m?2 (May) to 81.2 mg m?2 (February) in Line-DW. Sediment chlorophyll a concentrations were also high during winter compared with other seasons ranging from 4.4 mg m?2 (November) to 73.2 mg m?2 (February) in Line-CH (Fig. 5B). No clear spatial variations were observed in Line-DW and Line- CH. The ratios of chlorophyll:pheopigment were relatively high during winter and low during spring (Fig. 5C) ranging from 1.16 (June) to 1.77 (September) in Line- DW. The ratios in Line-CH were also high during winter but low during summer (Fig. 5D) ranging from 1.07 (June) to 2.3 (November). In general, the ratios, indirect index for grazing activity or poor growth environments were low, suggesting that benthic microalgae experience active grazing or environmental stress in the sampling sites.
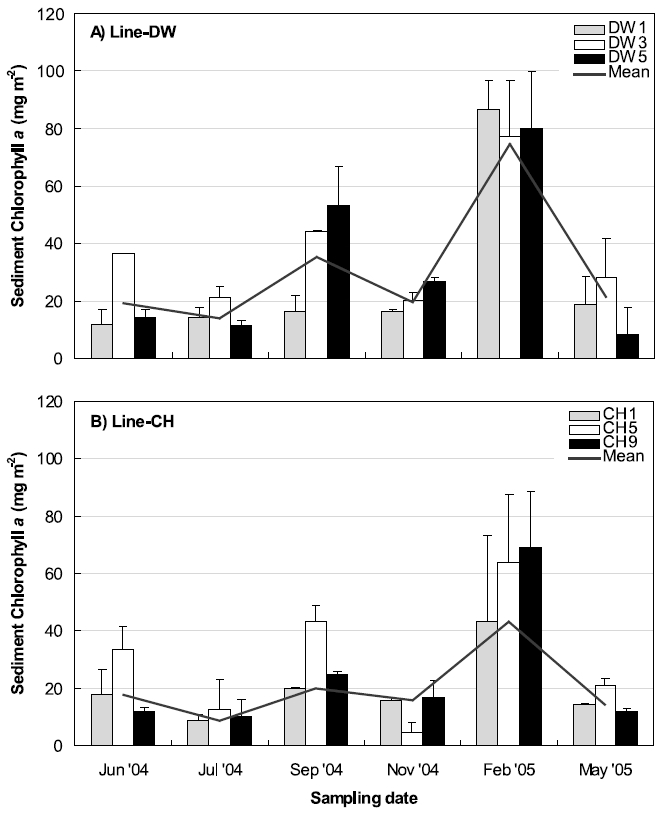
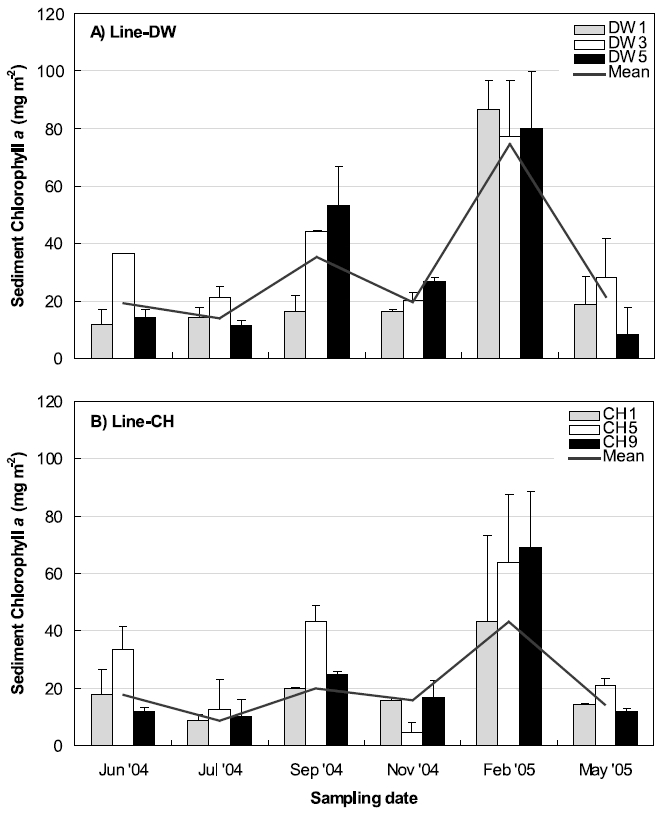
Three stations were selected between land boundary and boundary of open ocean at Line-DW and Line-CH respectively to compare the sediment chlorophyll a concentrations during the sampling periods (Fig. 6). Chlorophyll a concentrations were highest in February, 2005 at the Line-DW and CH whereas the pattern of spatial variations was reversed. Station close to land (DW 1) had higher concentrations than stations close to open ocean (DW3, 5) in the Line-DW while chlorophyll a were lower at the station landward (CH1) than stations seaward (CH5, 9). Minimum chlorophyll a concentrations were observed during July, 2004 in the Line-DW whereas minimum was detected during November, 2005 in the Line-CH especially at Station 5. In September (2005) relatively high chlorophyll a developed especially at the stations DW5 and CH5 and stations seaward had higher level than stations near land.
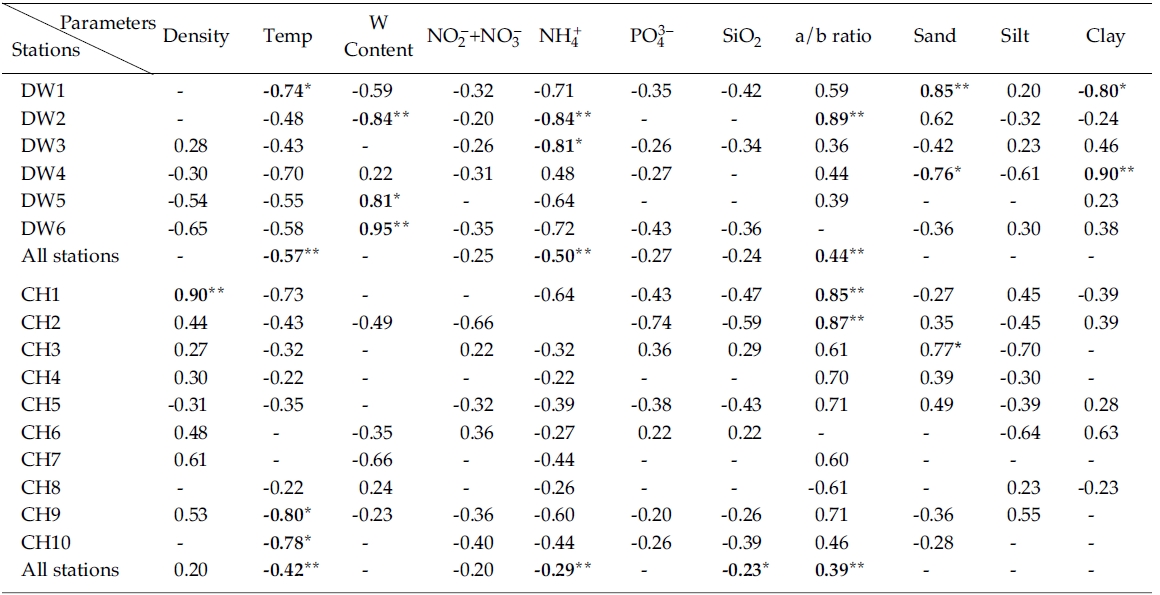
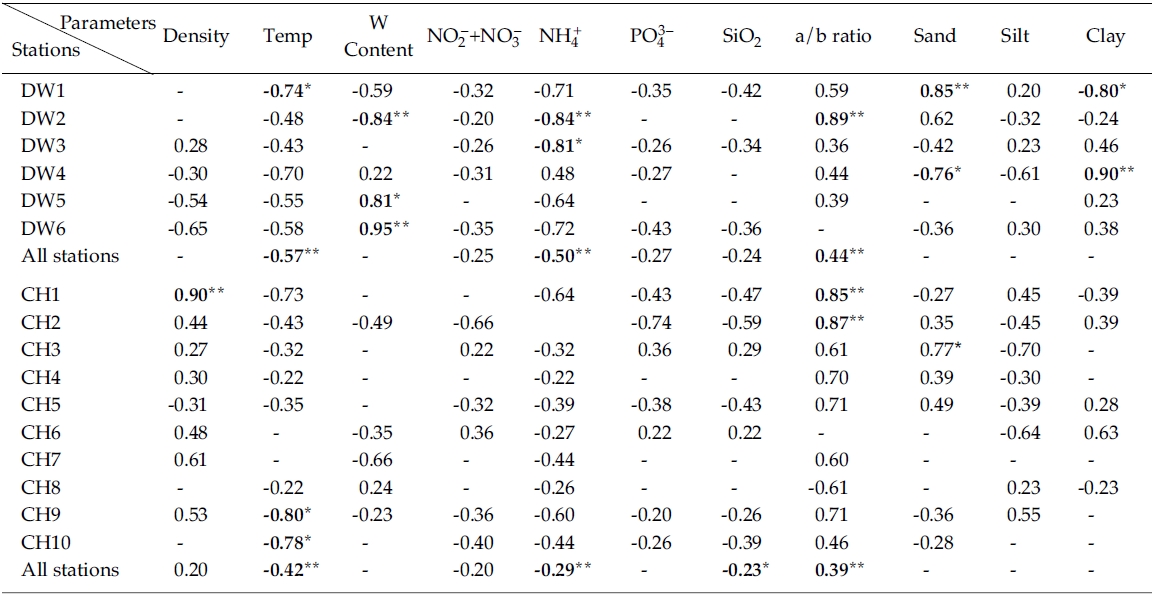
Results from simple linear regression analysis on chlorophyll a versus various parameters were shown in Table 1. Sediment bulk density was negatively correlated with chlorophyll a at stations seaward in the Line-DW but positively correlated in many stations the Line-CH. Sediment temperature had a negative relationship with chlorophyll a in the Line-DW and CH. Water contents in sediments were negatively correlated with chlorophyll a at station close to land (DW2) but positively correlated at stations near open ocean (DW5, DW6) in the Line-DW. Significant correlation between water contents and chlorophyll a was not observed in the Line-CH.
Negative correlation was observed between nitrate + nitrate and in Line-DW and similar relationship was generally observed in Line-CH except Stations CH3 and CH6 where positive correlation was detected. Significant negative correlation was observed between ammonium and chlorophyll a in the Line-DW and similar relationship was detected in the Line-CH. Orthophosphates and silicates were negatively correlated with chlorophyll a in Line-DW. In Line-CH chlorophyll a were generally negatively correlated with the nutrients except the Stations CH3, CH6 where positive relationship was observed.
The ratios of chlorophyll:pheopigment were positively correlated with in Line-DW and similar relation was detected in Line-CH except Station CH8, suggesting that benthic microalgae were influenced by physical environmental stress and/or biological interaction such as grazing. Percentage of sand in sediment texture was significantly and positively correlated with chlorophyll a at Station DW1 of Line-DW whereas percentage of clay was significantly and negatively correlated in the station. The relationship was reversed in Station DW4. In Line-CH, sand and clay were mostly positively correlated with chlorophyll a especially at stations near land whereas negative relationship was detected between silt and chlorophyll a.
Multiple regression analyses indicated that sediment temperature, water contents, chlorophyll:pheopigment, clay % and ammonium were responsible for 64% of the variation in sediment chlorophyll a in Line-DW. The sediment temperature, chlorophyll:pheopigment and nitrite+nitrate were responsible for 41% of the variation in sediment chlorophyll a in Line-CH.
Sediment microalgae play an important role both as food for benthic organisms and a source of nutrients for the overlying water column after decomposition (Middlelburg et al. 1996; Herman et al. 2000; Cook et al. 2004b). The importance of sediments as a source of ammonium to support water column primary productivity during the summer was documented for estuarine systems (Seitzinger and Nixon 1985; Nixon 1987; Pinckney and Zingmark 1993; Cowan and Boynton 1996 Cowan et al. 1996; Sullivan and Currin 2000). Nutrient concentrations in porewater of sediments in this study are greatly higher than those in water column near the study sites (c.f., Lee et al. 2001), suggesting that nutrient released from sediment are supplied to water column in the Kwangyang Bay. Trophic importance of the sediment microalgae to Kwangyang Bay food webs was reported by Kang et al. (2001, 2003). Biomass of sediment microalgae (chlorophyll a) ranged from 4.4 to 81.2 mg m?2 in this study and was relatively high compared with other tidal flat systems such as Mangyung (0.24-32.11 mg m?2) and Ganghwa tidal flat (1.18-34.25 mg m?2) in Korea (Oh et al. 2004; Yoo and Choi 2005). Primary production in sediments was generally high during spring or summer (MacIntyre et al. 1996). Biomass in this study was high during winter (February, 2005) and low during summer (July, 2004) as presented in Figs 3 and 4. The similar pattern of seasonal variations was observed in the Mangyung and Ganghwa tidal flat and diatoms were dominant in the microalgal population of the tidal flats (Oh et al. 2004; Yoo and Choi 2005). Earlier studies (Yoder 1979; Smayda 1980) documented that growth of diatoms in water column such as Skeletonema costatum was preferred by low water temperature and light intensity. The relationship between sediment temperature and chlorophyll a concentrations was reported from the comparison between field study and mesocosm experiments for a lagoon system in the Western Australia (Kendrick et al. 1998). Results from regression analyses in this study (Table 1) also showed that chlorophyll a concentrations were significantly negatively correlated with sediment temperature, suggesting that growth of benthic microalgae microalgae dominated by diatoms is affected by sediment temperature. High intensity of solar radiation during summer (Fig. 1) also may influence growth of benthic microalgae in the study sites since the samples were taken when the sediments were exposed to air. This hypothesis is supported by the result that chlorophyll a concentrations collected in Line-DW were significantly and negatively correlated with solar radiation in air from the correlation analyses in this study (R = ?0.33, p < 0.1). However, further studies including mesocosm experiments are required for better understanding the direct effects of temperature and solar radiation on sediment chlorophyll a in the tidal flat of Kwangyang Bay.
de Jonge (1985) reported that the diatoms were dominant in mud coating sand grains since mud rather than in the bare parts of the sand grains systems since mud serves as a substratum for diatoms on the tidal flats. Lower rates of primary production were observed at coarse grain-sized sediments (greater exposure to water energy) than fine grain-sized sediments (Cook et al. 2004a). Herman et al. (2001) proposed that high mud content in tidal flats reduce grazing pressure on microalgae from the ECOFLAT studies. In this study sand percentage in grain size of Line-DW was increased as seaward (Fig. 3G) but chlorophyll a concentrations were not varied (Fig. 5A). Results from regression analyses also did not show clear relationship between grain size and chlorophyll a concentrations (Table 1), suggesting that grain size was not major factor controlling the biomass of benthic microalgae in the study sites. In Line-CH, sand percentage in grain size extraordinarily increased at Station CH9 (Fig. 3H) whereas chlorophyll a concentrations were not higher than at Station CH10 (Fig. 5B) where sand percentage was low. The high percentage of coarse grain (sand) at Station CH9 is due to dumping sands into the sites for aquaculturing shot-neck clams (Ruditapes philippinarum).
Nutrient concentrations in the Line-CH were generally higher than those in Line-DW (Fig. 4). This difference may be resulted from different water contents (Figs 3C and 3D) caused by gradient of grain size in sediment between the Line-DW and Line-CH (Figs 3G and 3H). A more stable habitat with higher concentrations of reduced nutrients such as ammonium has been reported to be favored by benthic microalgae within soft sediments (Saburova and Polikarpov 2003). The preference of microalgae to the stable habitat with high levels of nutrients was not observed in this study since chlorophyll a concentrations were not significantly different between the Line-DW and Line-CH in this study (Figs 5 and 6). However, significant negative correlation of ammonium vs. chlorophyll a concentrations in sediments (Table 1) suggests that benthic microalgae prefer ammonium, more reduced N to nitrite+nitrate and ammonium availability may affect the growth of benthic microalgae in the study sites.
Aquatic and benthic microalgae biomass is controlled by top-down such as grazing by consumers as well as bottom-up mechanisms including physical and chemical variability (Goldfinch and Carman 2000; Worm et al. 2002; Pinckney et al. 2003). In this study, quantitative relationships between meiofaunal grazers and microalgae were not investigated but the ratio of chlorophyll a and pheopigments was used to examine the grazing effects. The ratios in the study sites were relatively low (1.41 ± 0.15 for Line-DW, 1.36 ± 0.19 for Line-CH), suggesting that grazing by meiofaunal consumers affects biomass of benthic microalgae. chlorophyll a concentrations were high during winter concomitant with high chlorophyll: pheopigment ratios (Fig. 5). Significant correlation of sediment Chlorophyll a concentrations vs. chlorophyll:pheopigments was also observed (Table 1). Tight coupling of trophic relationships between macrobenthosand microalgae reported by Kang at al. (2003) for the intertidal flats of Kwangyang Bay also support the scenario of bottom-up controls on benthic microalgae in the study sites. However, pheopigments can also be produced within phytoplankton cells during senescence caused by poor growth environments or prolonged exposure to the dark (Yentsch 1967; Daley and Brown 1973). Thus, it is difficult to distinguish the relative importance of grazing vs. other factors related to bottomup control in regulating the phytoplankton community without the direct investigation on grazing effects in the study sites.
Algal colonized sediments can utilize nitrate, and thus reduce nitrogen fluxes to the water column (Tyler et al. 2003; Sundback et al. 2004; Sin et al. 2006). In addition oxygen produced by microalgal photosynthesis stimulates nitrification (an aerobic process), thus inducing coupled nitrification-denitrification (Risgaard-Petersen et al. 1994). At low nitrate concentrations, however, nitrification- denitrification may be suppressed by microalgae due to microalgal utilization of the nitrate at the sediment- water interface (Sundback et al. 2004). Results from the regression analysis of this study showed that nitrite + nitrate concentrations in porewater were negatively related with sediment chlorophyll a concentrations but the relationship was not significant (Table 1) although nitrite+nitrate concentrations were greatly higher especially during summer and fall than those in water column near the study sites (c.f., Lee et al. 2001). Instead, significant negative correlation of ammonium vs. chlorophyll a concentrations was observed, suggesting that benthic microalgal community reduce benthic ammonium fluxes from sediments to water column. Nitrite + nitrate and orthophosphate concentrations were high during summer and fall but rapidly decreased during winter when chlorophyll a concentrations were high, suggesting that benthic microalgae utilize the nutrients during winter. Silicate concentrations were also significantly and negatively correlated with chlorophyll a and the results support the feature that benthic microalgae reduce benthic nutrient fluxes especially for ammonium and silicates from sediment to water column in the intertidal flats. The porewater nutrients at high concentrations during summer and fall may be diffused into water column and contribute to phytoplankton blooms which often develop during summer in water columns of the Kwangyang Bay (c.f. Kwon et al. 2004).