The spatial aggregation of species along a gradient is termed zonation (Smith and Huston 1989). It has been demonstrated that environmental heterogeneity affects plant distribution due to variations in micro-environmental conditions, and the specific responses of plants to this variability (Ranwell 1972, McCook 1994, Culter et al. 2008). Several studies have investigated the relationship between vegetation and environmental gradients (Lortie and Cushman 2007, Lane et al. 2008, Carboni et al. 2011). Variation in vegetation landward across coastal sand dunes is associated with decreases in the degree of exposure to salt spray (Lubke 1983, Maun and Perumal 1999), strong winds, and sandblast (Martinez et al. 2001) as well as with increased nutrient levels (van der Valk 1974, Gilbert et al. 2008, Lane et al. 2008 ) and moisture content of the developing dune soils (Baldwin and Maun 1983). Variation in soil properties may also result from topographic heterogeneity. The resultant soil-vegetation and soil-landscape interrelationships should therefore be expected to be more complex than either of the two considered separately (Chen et al. 1997).
Many natural sand dune areas in South Korea have been lost due to development, overuse, invasive nonindigenous plants, and beach erosion by typhoon surge, coastal flooding during the summer, aggravation of the meteorological disaster and sea level arising (Yoon et al. 2010, Kim et al. 2013), resulting in beach regression, biodiversity loss, damage to vegetation, and environmental disturbance which in turn have affected the re-vegetative process (Girad et al. 2002). In coastal ecosystems, these impacts include the interruption of the sand supply, which can accelerate sand erosion, and result in the deterioration of the dune system. In addition to the collapse of a balance of eroded and deposited sand by foundation of shore protection structures is as an important factor influencing sand erosion. Although the importance of maintaining beach and dune ecosystems is increasingly recognized (Mendoza-González et al. 2013) and many studies on coastal dune protection have conducted, there is still insufficient ecological information on dune vegetation and environmental factors.
The objectives of this study were therefore to describe the distribution patterns of coastal sand dune plants and the soil environment across the dune gradient and to relate plant distribution with environmental factors.
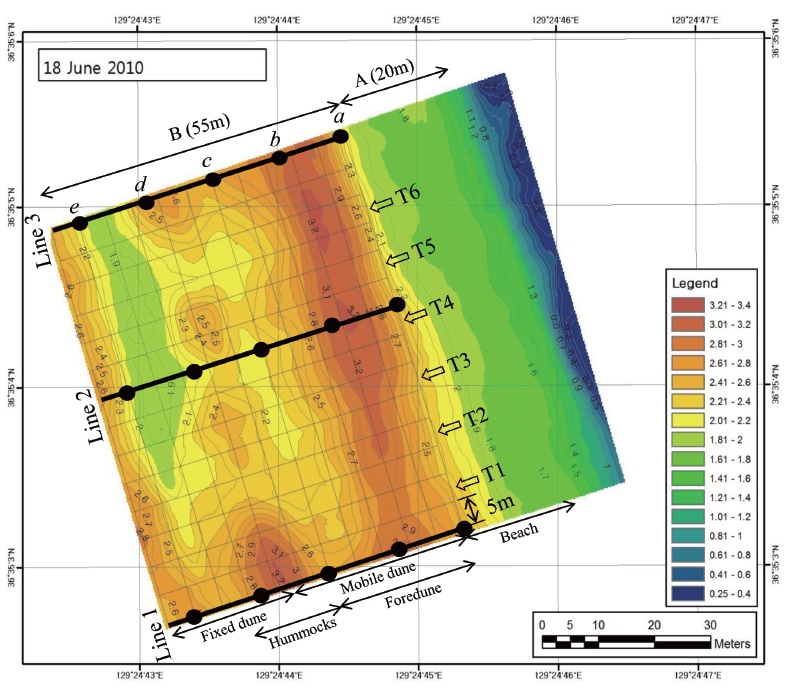
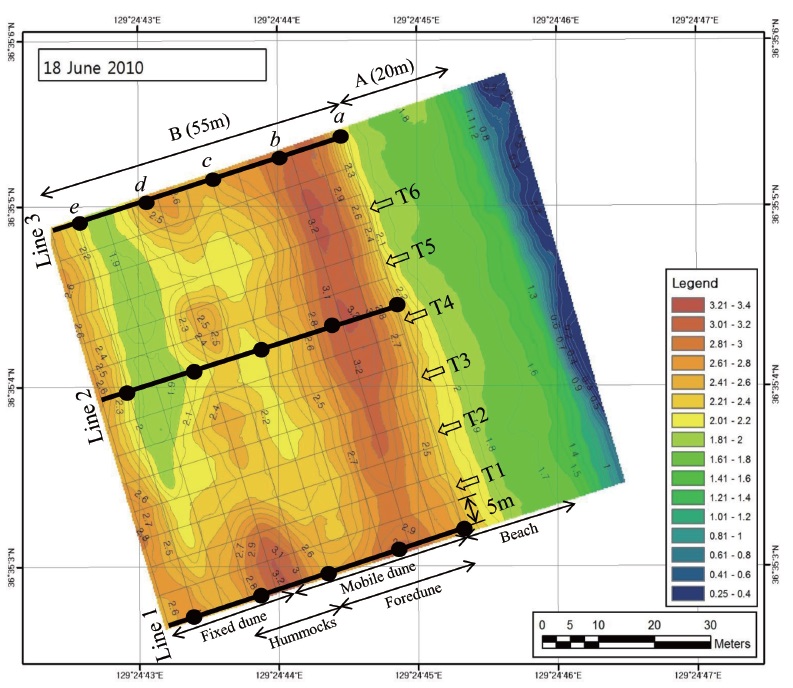
Even though a lot of coastal dune areas in east coast have been affected by natural or artificial environmental conditions, the Goraebul coastal dune field, the largest that in east coast of Gyeongsangbuk-do Province in Korea (36°34′27.9″N, 129°25′04.6″E), has well-developed foredunes and dune vegetation and is an established Long-Term Ecological Research (LTER) site for the study of coastal ecosystems (Fig. 1). Since it located with an area managed by the military, the Goraebul coastal sand dune system is relatively unaffected by anthropogenic activities compared with the surrounding region due to restrictions on development and limited accessibility from the outside. The sand deposition in almost seasons except summer was investigated in Goraebul sand beach (Bang and Lee 2011). The microtopography includes a beach face, berm, beach flat, foredune and three hummock dunes lying parallel to the shoreline (Fig. 1). Sandy beach significantly influenced by wave action was developed about 20 m in width and was rarely vegetated.
Temperature in this region fluctuates seasonally; the warmest and coldest months are August and January, respectively. Mean annual temperature and precipitation for the study period (2010-2013) were obtained from meteorological records of the Yeongdeock Meteorological Observatory. The mean annual temperature is 13.2℃ (January 0.0℃ , August 26.4℃). The mean annual precipitation is 1000 mm, of which 670-900 mm fall in April-October. The mean monthly temperature in April-September varied between 10.3-15.5℃. Extreme mean monthly temperatures ranged between -2.6℃ (January 2011) and 27.7℃ (August 2013).
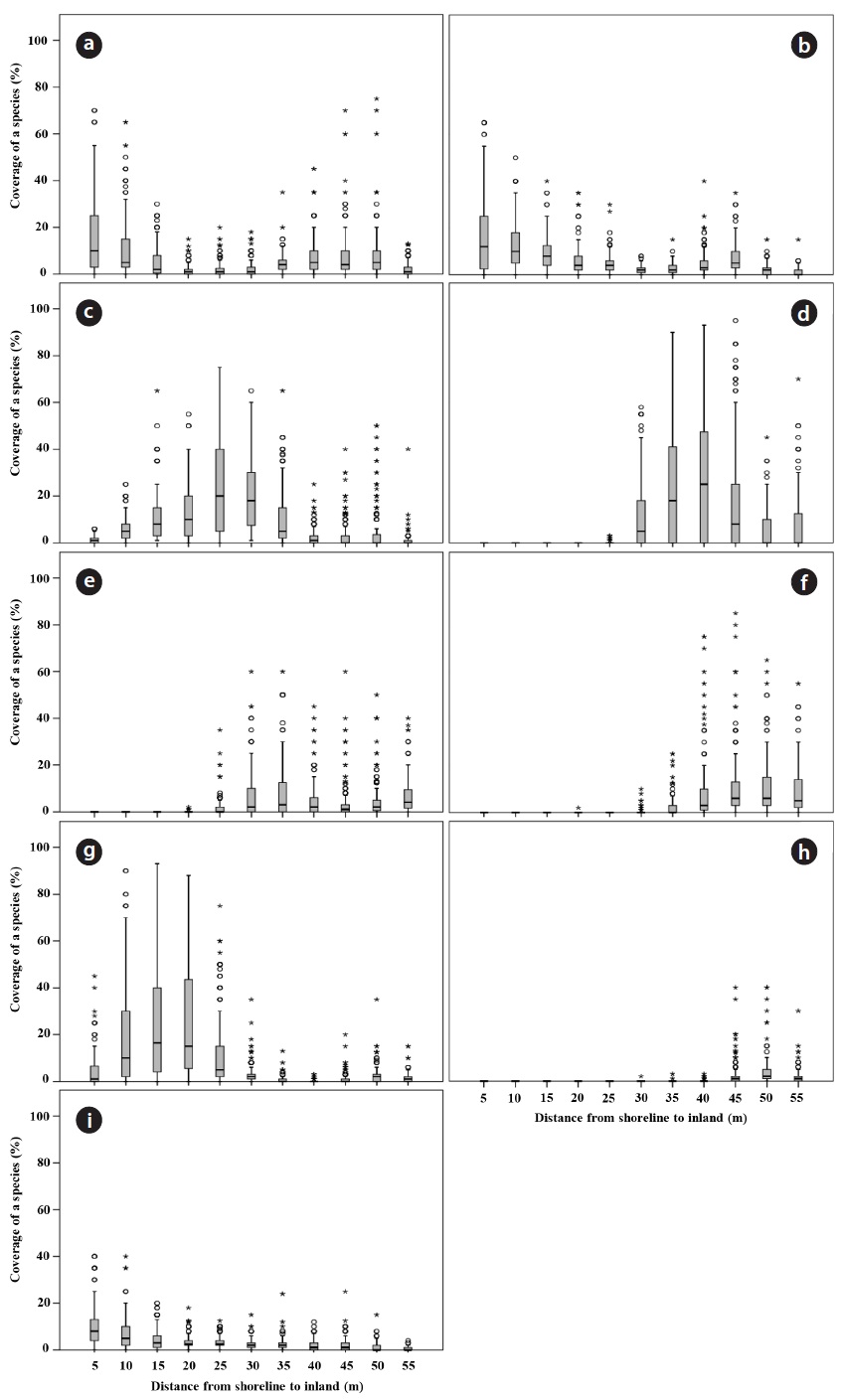
To reveal the distribution patterns of plants in the Goraebul coastal sand dune, a rectangle of 70 m × 55 m containing 154 grids of 5 m × 5 m grids which extended from the location of the first plant found on the upper shore to the fixed dunes was established (Fig. 1). The surveying first line which was established parallel to the shoreline was set at the distance of about 20 m from the shoreline or about 10 m from the berm. To identify spatial patterns of plant species across the gradient, we recorded coverage of all plant species in a grid that consisted of six belt transects, each 55 m long, which were separated by a distance of 5 m (Fig. 1). Plant cover was surveyed over a four year period (from April to October 2010-2013). Within each quadrat, the percent cover of plant species was first assigned to one of the six cover-classes: 0-5%, 5-25%, 25-50%, 50-75%, 75-95%, 95-100%, then plant cover was further refined to the nearest 5% for cover values from 5% to 95% and to the nearest 1% for cover less than 5% and greater than 90%.
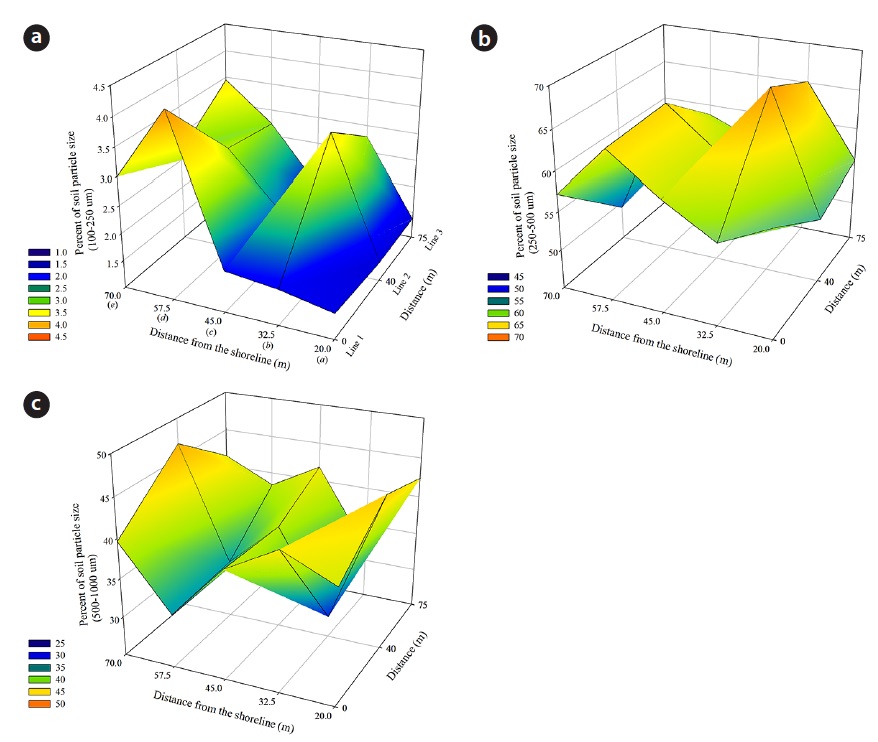
In the study area, five quadrats (2 m × 2 m) were established at each of the three lines to analyze the relationship between distribution of plants and environmental variables, thus producing data from 15 quadrats (Fig. 1). Soil samples (0.2 kg) were collected from 0 to 10 cm depth at each the 15 quadrats during the period April - October from 2010 to 2013. Water content (WC) of the soil was determined by comparing the weight difference between fresh and oven-dried soils. Soil particle sizes (SPS) were analyzed by a Particle Size Analyzer (Mastersizer 2000; Malvern Instruments Ltd, Worcestershire, UK) which allows particles in the size range 0.02 to 2000 μm, and the soil had a particle size distribution of 99% and more sand (150 to 1000 μm), classifying this soil texture as sand (United States Department of Agriculture 1987). The sand separate was subdivided into coarse sand (500-1000 μm), medium sand (250-500 μm), and fine sand (100-250 μm).
Soil samples (5 g) were added to 25 mL distilled water, and shaken for 1 h. The soil solution was then filtered through a filter paper (Whatman No. 40, 110 mm), and the soil pH and total ionic content (TIC, calculated as NaCl equivalents) were measured using a pH meter (Orion US/710; Thermo Orion, Waltham, MA, USA) and an electrical conductivity meter (MX300 X-masterpro; Mettler-Toledo, Columbus, OH, USA), respectively. The organic matter (OM) contents were estimated by loss of dry mass upon ignition at 550℃. Total nitrogen (TN) was measured by the Kjeldahl method.
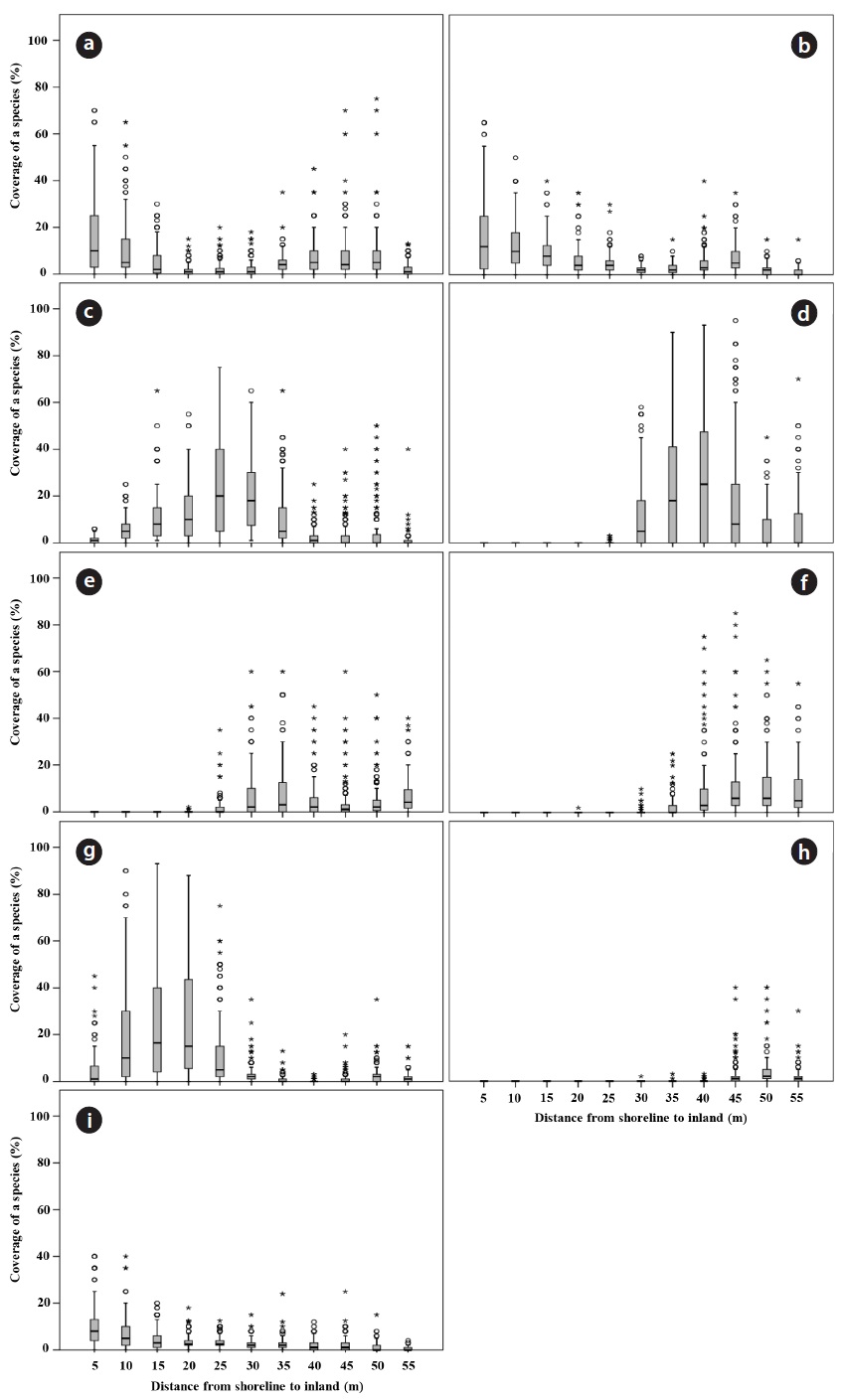
Box-whisker plots were, generated using SPSS ver. 18.0 (SPSS Inc., Chicago, IL, USA), and used to illustrate the plant distribution at varying distance from the shoreline during the study period. The spatial distribution pattern of soil parameters was illustrated using Sigma-plot 2000 (ver. 6.10; SPSS). The data were tested for normality with the Kolmogorov-Smirnov test and homogeneity of variances with Levene test. To test the relationship between the distance from the shoreline and soil properties, Spear-man’s rank correlation was applied due to lack of homogeneity of most soil variables except TN. Quantitative data on soil properties among soil sample plots as factor were compared using parametric (ANOVA, followed by Duncan’s test as a post hoc analysis) or non-parametric analysis of variance (Kruskal-Wallis test when the data were not normally distributed or the variances of the groups differed significantly, followed by Mann-Whitney
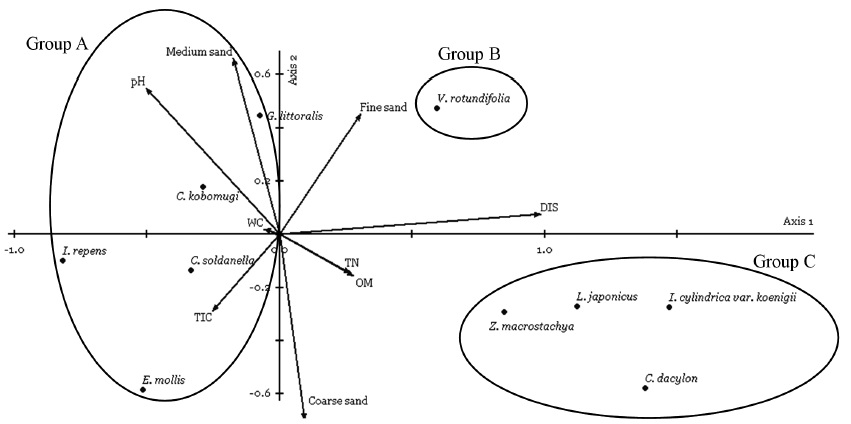
In order to obtain an effective description for sand dune vegetation and related environmental factors, both two-way indicator species analysis (TWINSPAN) and canonical correspondence analysis (CCA) are employed. CCA is used to identify the most important environmental factors in determining patterns of species distribution, leading to an ecologically meaningful interpretation of the essential gradients in a few dimensions (Palmer 1993). Both vegetation classification and ordination analysis were performed on a matrix containing ordinal transform values for all species according to van der Maarel (1979) replacing the modern nine-point Braun-Blanquet scale by the values 1-9. 13 species of the total flora occurring only at a single sample plot are eliminated from the species data set prior to TWINSPAN and CCA using PC-ORD program for windows version 5.0 (McCune and Mefford 1999). Therefore, a matrix of 10 species × 323 releves resulting from trnansect sampling was subjected to TWINSPAN and CCA. Eight soil parameters (WC, pH, TN, OM, TIC, fine sand, medium sand, coarse sand) and distance from shoreline were included for CCA. The following options for CCA were used in PC-ORD: row and column scores were standardized by centering and normalizing; ordination scores were scaled to optimize rows; site scores were linear combinations of variables, and a Monte Carlo test of significance was used with 9,999 randomizations.
>
The spatial distribution of plants
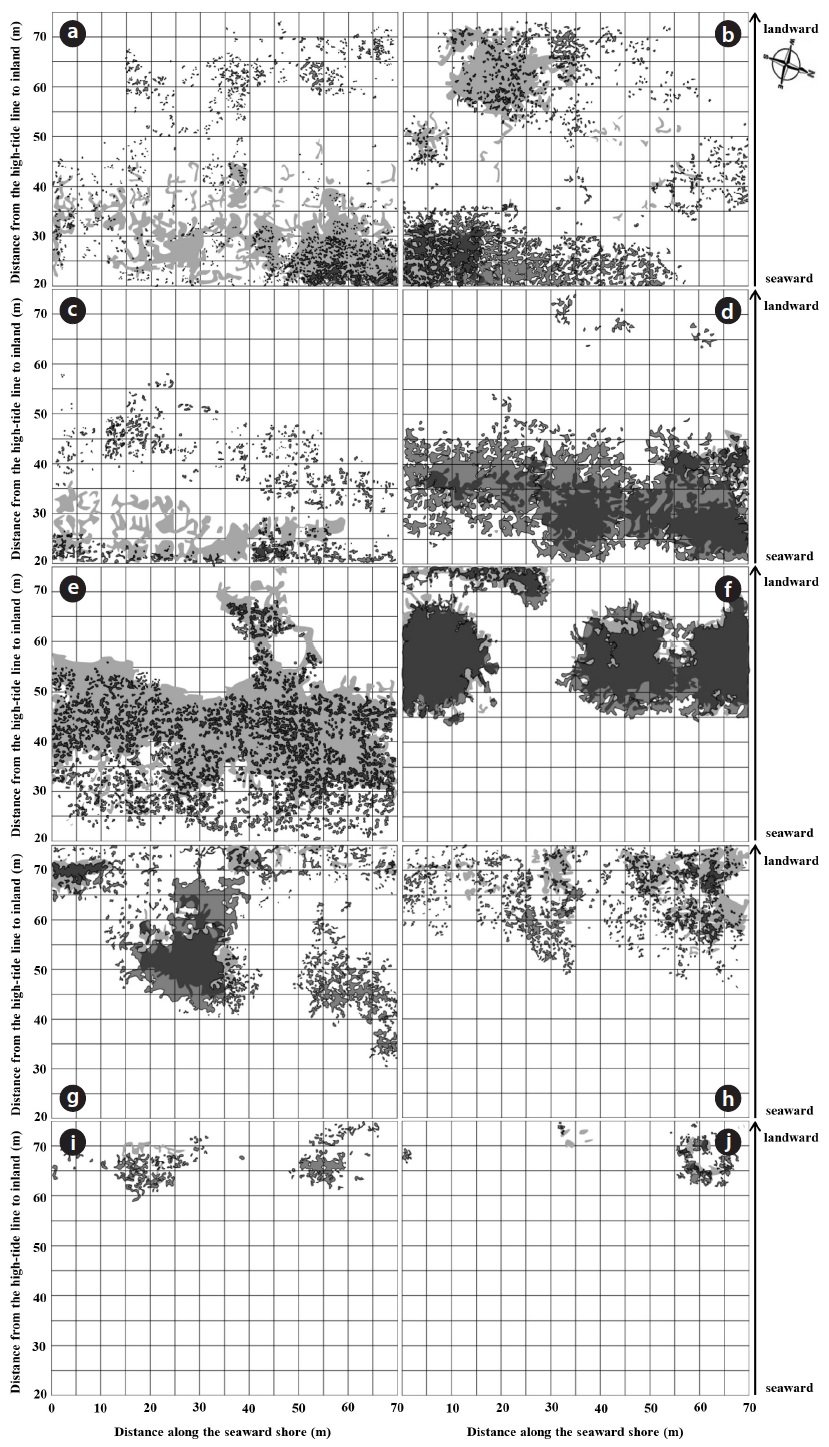
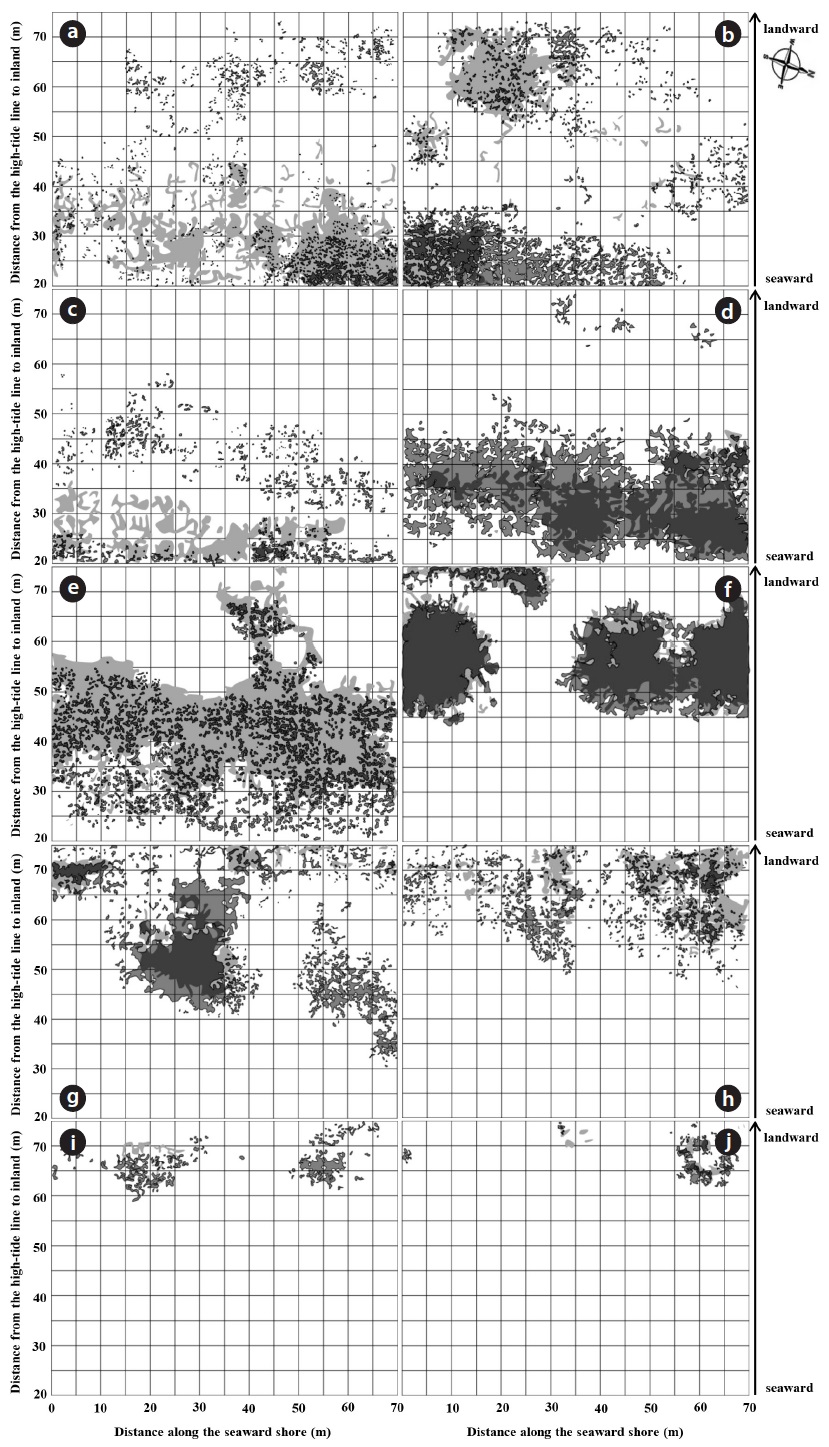
A total of 23 species and 12 families were recorded in study site during the four year period. Dicotyledons comprised 65.2% of the total (15 species in 9 families), while the remainder consisted of 7 monocotyledons species (2 families) and one gymnosperm. The Gramineae family exhibited the highest number with 6 species, followed by Asteraceae, Chenopodiaceae and Convolvulaceae with 4, 3 and 2 species, respectively. Coastal sand dune plants constituted 68.2% of the recorded species. In terms of life duration, 13 species were perennials (56.5%), 8 species were annuals (34.8%) and 2 species were biennials (8.7%). The number of plant species increased with increasing distance from the shoreline. The highest diversity was observed on the relatively protected backdune. The number of plant species per quadrat was lowest at 20 to 25 m from the shoreline (mean 4.11 ± 0.41), and highest at 65 to 70 m (mean 7.40 ± 1.09). Change in the species composition in a seaward to landward direction was also evident (Fig. 2). The change of plant coverage along the distance from shoreline to inland during the four years was appeared using box-whisker plot from SPSS 18.0 because of seasonal fluctuation in plant cover (Fig. 3).
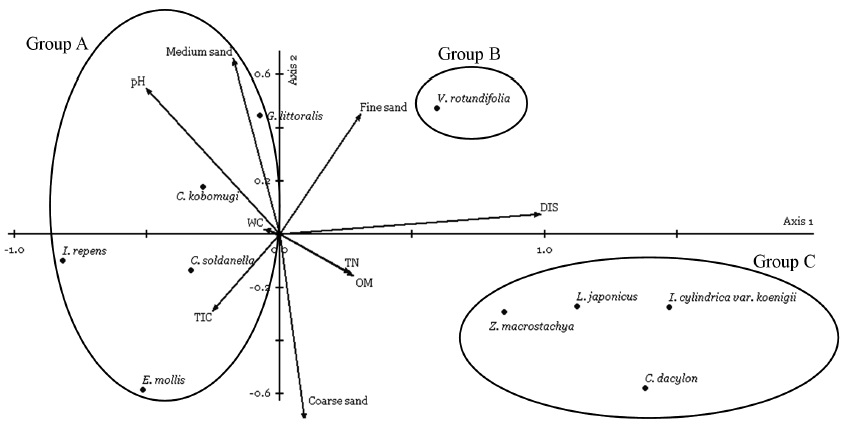
The application of TWINSPAN classification based on the coverage of 10 plant species recorded in present study site over four years led to the recognition of three vegetation groups that are associated with the following environments: foredune, semi-stable zone (hummocks), and stable zone.
Group A. This group was characterized by
Group B. This group was characterized by
Group C. Indicator species of this group were
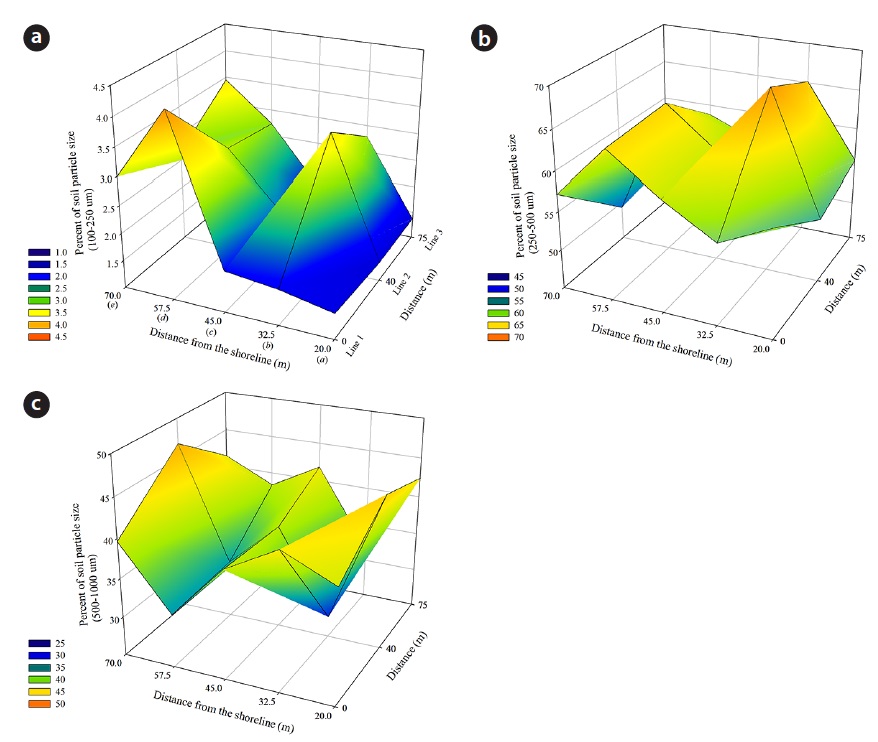
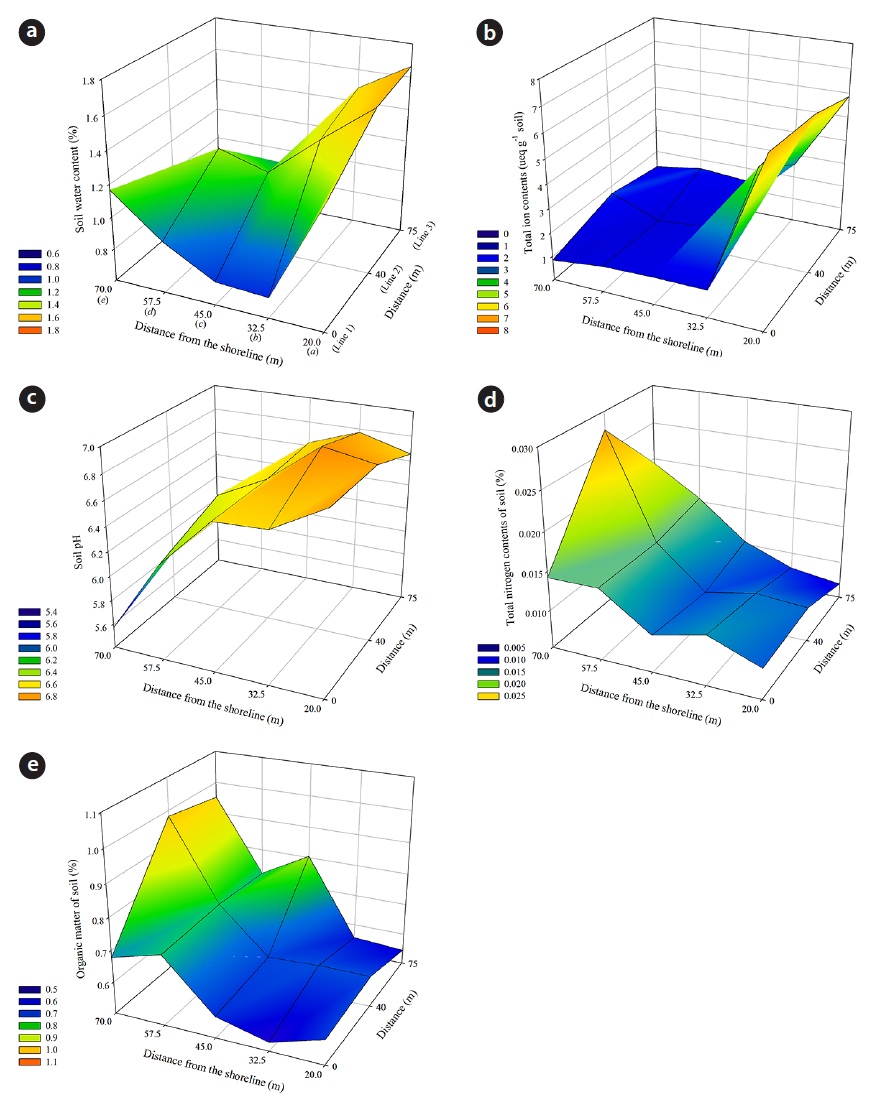
The size of sand grains generally ranged from 160 μm to 1400 μm. The percent composition of soil particles at the 18 quadrats of the study site, as measured from April 2010 to October 2013, was divided into three size classes: 2.43 ± 0.91% of the sand particles were fine (100 to 250 μm); 58.63 ± 5.06% were medium (250 to 500 μm); and 38.55 ± 5.37% were coarse (500 to 1000 μm). These data indicate that the Goraebul coast primarily comprises relatively medium-textured material, as the highest proportion of soils were medium in texture, and consisted of particle sizes between 420 to 550 μm, The spatial distribution of the mean percent soil particle size during the study period is shown in Fig. 4. The proportion of sand grain size also varied among habitats of dominant species. A relatively high porportion of fine sand was shown at Line 1
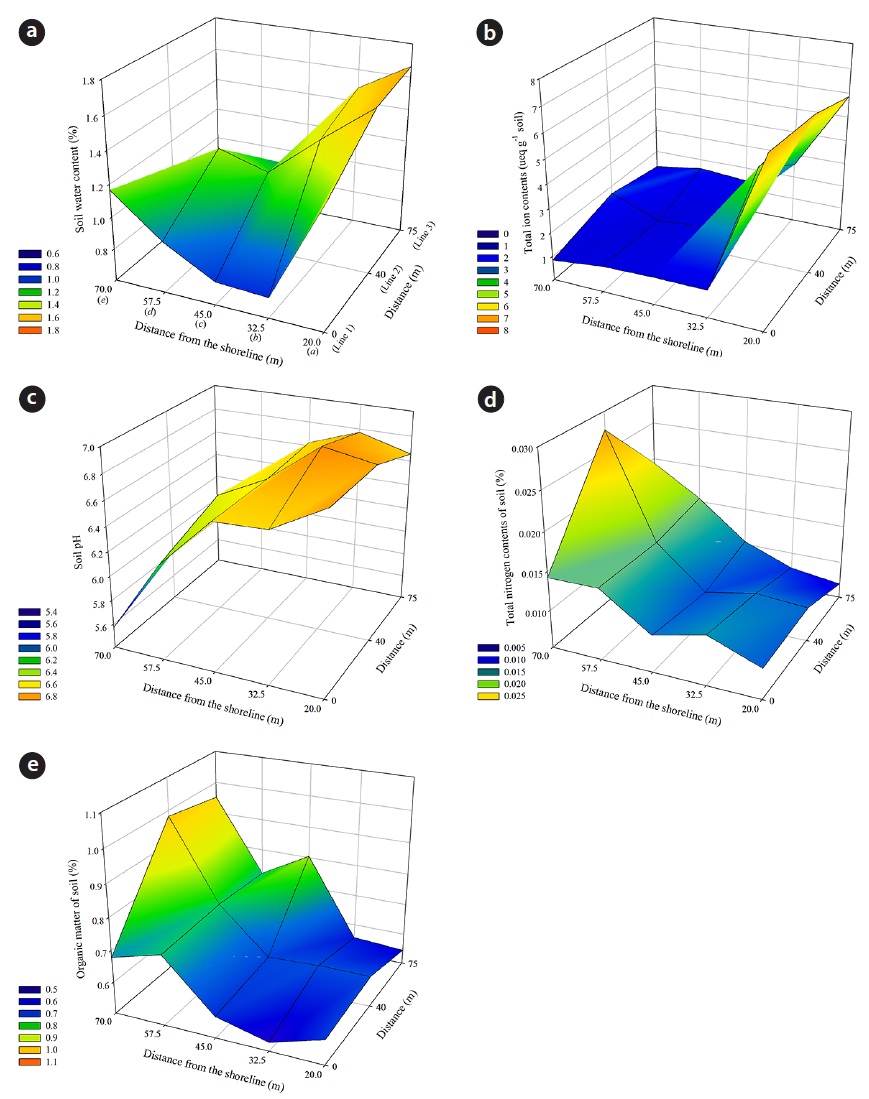
Soil WC, pH and TIC showed a negative correlation to the distance (
TIC were decreased with increasing distance from the shoreline, and a significant difference between soil sample plots was found (Kruskal–Wallis test:
TN increased with distance from the shoreline, and the highest values were observed in Line 2
>
Vegetation-Soil Relationships
CCA ordination was performed using PC-ORD to identify which soil environmental gradients determined the distribution patterns of coastal sand dune plants (Fig. 6). The CCA eigenvalues for the first two ordination axes were 0.397 and 0.117, which explained 23.4% of the variance in the community data. The interset correlations of soil environmental factors with axis scores showed that distance from the shoreline had a higher correlation with first axis, followed by OM, TN, and TIC. Soil pH and fine sand had relatively high correlation with first and second axis. Cor-relation with medium and coarse sand was the significant soil property along the second axis. Soil WC showed the lowest correlation with first and second axis.
Coastal sand dune species occurred on the landward such as
The phytosocialogical study on plant species distribution of the East Coast in Korea has been described in other studies (Jung and Kim 1998, Min and Je 2002, Kim 2005, Han et al. 2013), and the patterns of species distribution in the site that we studied was similar with it described in earlier studies. Kim (2005) classified communities on sand dunes by TWINSPAN, CCA, and detrended canonical decorrespondence analysis (DCCA):
Understanding the mechanisms driving community assembly and shifts in plant distribution as species expand in newly favorable areas and decline in increasingly hostile locations has been an important focus in plant ecology. In the present study, the distribution of individual species changed from 2010 to 2013 (Fig. 2). The change of plant distribution is related to various variables such as edaphic conditions, climate and biotic interactions (Wisz et al. 2013, Beauregard and de Blois 2014), which could limit plant species’ ability to establish in areas newly within their niche. Species co-existence within communities, and species sorting along environmental gradients, are thought to be regulated by the degree to which species share similar adaptations (Kelly and Goulden 2008, Carboni et al. 2013).
The texture of the sand on the sand dune ridges is typically finer than that of the beach and slack (Maun 2009). In this study, the highest percentage of soil particles of the size fraction 100-250 μm were found on hummuck dominated by
Ruocco et al. (2014) suggested that a distance to the coastline, mean grain-size, shoreline trend and OM as abiotic factors were found to be closely correlated with the distribution and composition of Mediterranean plant dune communities. Environmental gradients were observed over the distance from the shoreline to inland areas at the Goraebul coastal sand dune study site. WC, pH and TIC decreased with increasing distance from the shoreline, but TN and OM increased. Although the value of soil WC in seaward slops of foredune was relatively high comparison with other sites, the results of variance analysis and CCA showed no significant differences between moisture contents at the soil sampling site and had a low score for any of the axis, respectively, indicating that soil moisture in this dune system could not explain the spatial distribution of coastal plants. In the study by Ishikawa et al. (1995), although soil WC did not show significant differences among different distance from the shoreline, the increase of water availability with increasing distance from the shoreline was caused by an increasing proportion of fine sand at inland sites beyond 60 m.
The distribution pattern of
TN was the highest in the
The increase of OM and nitrogen contents in a stabilizing habitat can facilitate the arrival or spread of other species, particularly invasive species already established in a coastal sand dune system, and the introduction of new non-native species into the region. At the study site, inland species such as
The spatial distribution of plants in the Goraebul coastal sand dune system may result from the interactions between the plant species and environmental heterogeneity. The spatial pattern of soil was found to be associated to vegetation types. Although environmental factors may act alone or in combination with other factors, distance from shoreline, soil pH, TIC, spatial variability in soil particle size, and the accumulation of OM and nitrogen was closely related to the spatial distribution of species in study area. The sand width is important in dune formation and the sand transport. The results of SPS proportion, particularly with respect to the proportion of fine sand size suggest that the morphology of a dune and the pattern of sand movement are closely related to the occurrence and diversity of plant species, their density, and community structure. In nutrient-poor sandy soil, especially stable back dune, mixed herbaceous community comprised of nitrogen fixing
The disruption of sand movement and beach erosion by construction and other anthropogenic activities in coastal sand dune could damage this coastal ecosystem. In order to maintain coastal ecosystem, it is therefore necessary to understanding of the controlling factors that are related to the distribution of various coastal sand dune plants. Sustainable management of dune system is imperative to maintaining its most beneficial environment and should take into account the positive effects of moderate sand movement.