Ecological restoration is defined as the process of assisting the recovery of an ecosystem that has been degraded, damaged or destroyed (Society for Ecological Restoration International Science & Policy Working Group 2004) and starts with the desire to improve such deteriorated landscapes or ecosystems (del Moral et al. 2007). In the past, most forest restoration projects aimed to rehabilitate abiotic functions and the Young-il soil erosion control project also aimed primarily to prevent the spread of secondary damage from forest soil erosion into aquatic, agricultural and human systems over the region. Recently, the project has been noted as successful landscape level restoration model to environmental managers and ecologists (Cho 2005, Lee and Suh 2005).
Forest landscape degraded can be rehabilitated to secondary forest through artificial modification such as plantation and soil erosion control facilities for deteriorated sites. So, landscape change or restoration exhibited by both the natural processes and the artificial assists including management practices and policies. Human dominated landscapes or region as like Korea, the artificial disturbance rules the change and configuration of the landscape (Cho 2009). As this point, the structural and compositional effect of introduced (
Partly, several studies tested differences in species composition and diversity between or among vegetation types, as Ito et al. (2004) on previous land use, plantation and secondary, Graae et al. (2003) on landscape context, isolated or not, and Graae and Heskjær (1997) on management history, unmanaged and managed. Synthetically results of those investigations revealed that seed disposal, seed bank and abiotic conditions, such as nutrients, soil water content, landscape context and so on, were attributed to the gap of vegetation types tested. For those results, there are fundamental differences on the starting point with absence and presence of initial floristics of vegetation development of Young-il restoration project and on the suffering active restoration treatment. Of particular important is the recognition that some ecosystems may occur in a number of alternative states, which may be contingent on the disturbance history, human intervention and so on (Beisner et al. 2003, Suding et al. 2004, Hobbs 2007).
Despite it has been mentioned that the successful restoration in landscape level was achieved in the Young-il district (Cho 2009), quantitative and functional evaluation of restored plant communities natural secondary (
This study area was conducted in plantation forests,
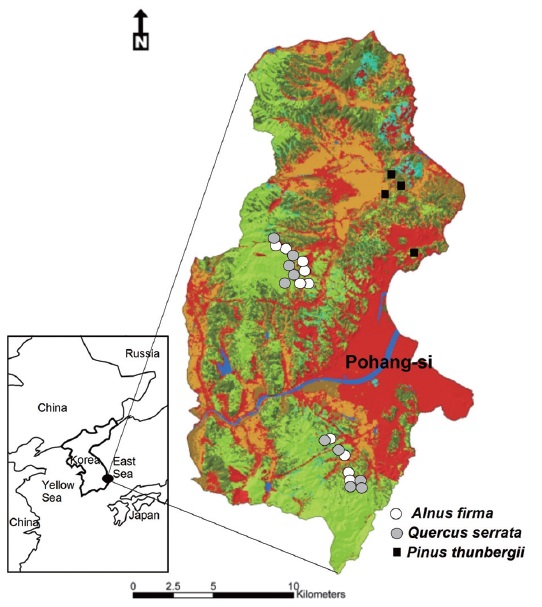
In the early of 1970s, remained natural vegetation of Young-il area was composed of
During the restoration project, various introduced, representatively
Hong (1982) and Cho (2005) reported that potential natural vegetation was oak dominated forest in lowland of Young-il erosion control district. Various plantations in study area were converted to oak dominate or co-dominate forest by natural succession (Cho 2005). While, for the
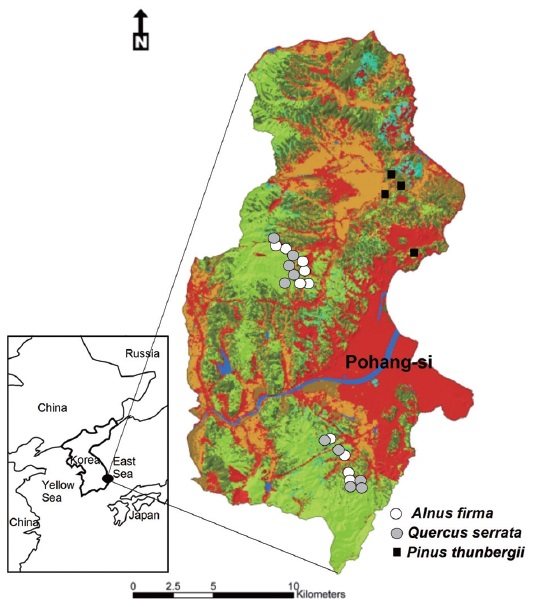
For restored forest, total of 14 stands were selected in
Vegetation sampling was conducted from 25 August to 12 September 2008 and 2009. Total of 24 with 10m × 50 m (500 m2) plots, a wide band for forest structure and composition of tree species, were established parallel with the contour and was > 30 m from all stand edges and road. The band consisted of 10 microplots (1 m2) for ground vegetation spaced at 5 m intervals, 5 subplots (25 m2) for shrub vegetation spaced at 5 m intervals and 2 ~ 5 plots (100 m2) for tree species composition. Within each sub (1.5 m ~ 2.5 m in height) and microplots (< 1.5 m in height), cover (%) of all vascular plants were recorded. Litter and coarse woody debris coverage (%) were also determined in microplots. Diameter at ground 0.3m height of all tree species was measured in a band. Age of each stand was estimated by counting tree rings from increment cores for the largest tree species in the plot. Mean age of
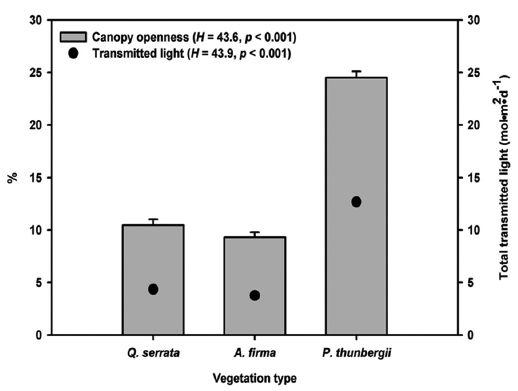
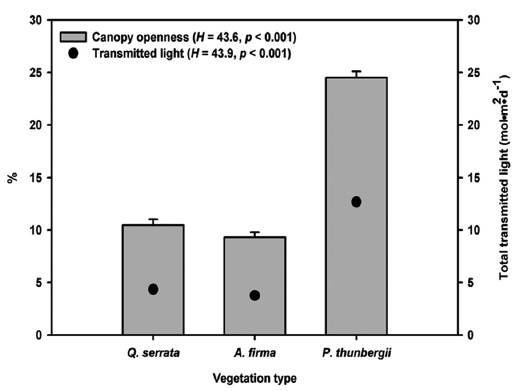
To estimate light availability and canopy openness (%), we took digital hemispherical photographs at the center and four corners of bands from a height of 1.8 m using Nikkon D80 digital camera with Sigma 4.5mm F2.8 EX DC Circular Fisheye HSM lens. Pictures were taken on uniformly overcast days to avoid direct sunlight. Total transmitted light or PPFD (Photosynthetic Photon Flux Density) (mol m-1 day-1) averaged over the year was calculated with Gap Light Analyzer 2.0 employing the standard overcast sky model (Frazer et al. 1999).
Stand-level means of
In addition to NMS, multiresponse permutation procedures (MRPP) (Biondini et al. 1988) to test whether species composition differed between natural and restored stands were applied. MRPP is a nonparametric procedure that avoids distributional assumptions of normality and homogeneity of variance, which are not commonly met in ecological data. It produces A-statistics (the chancecorrected within-group agreement) with probability of statistical significance (McCune et al. 2002).
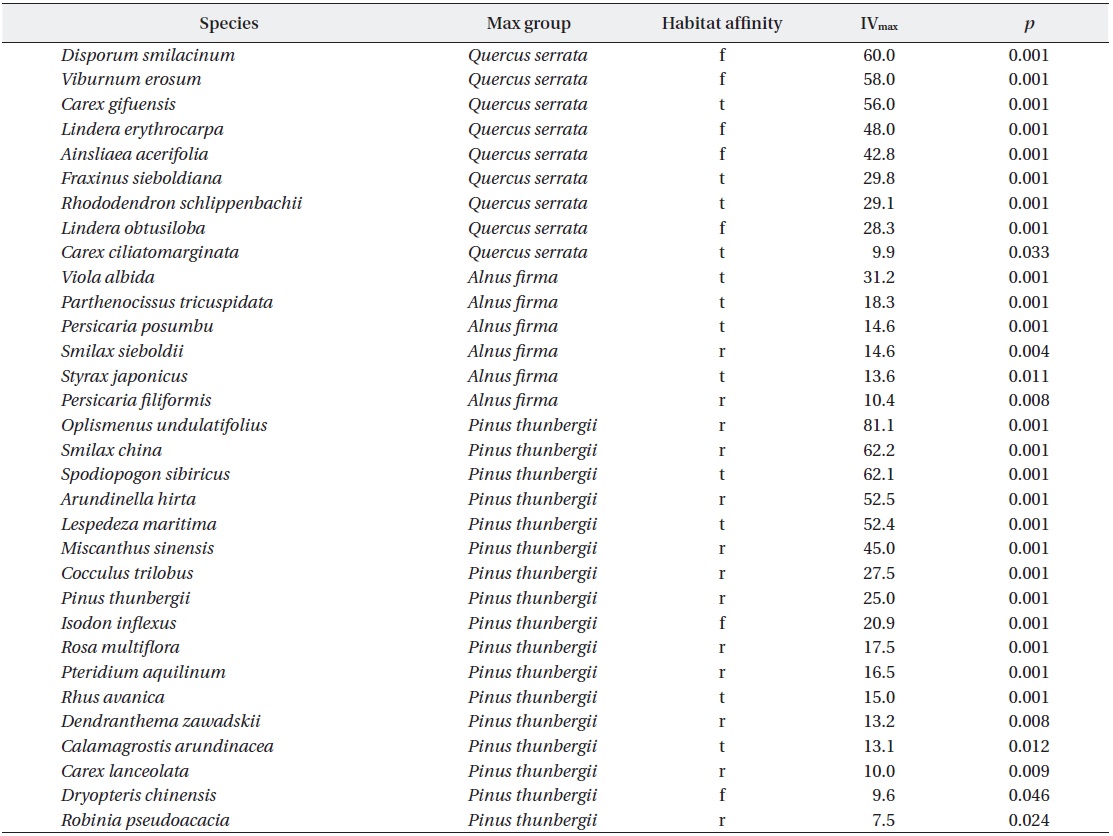
For compare species attributes as habitat affinity, indicator species for each stands were identified with ISPAN (indicator species analysis; Dufrêne and Legendre 1997, McCune and Mefford 1999). The indicator values (IV) as a result of ISAPN range from 0 to 100 and 100 as perfect indication means that presence of a species points to a particular group without error (McCune et al. 2002). For yielded indicator species, habitat affinity of forest, transitional and ruderal was classified by applying field professionality of the authors. Kruskal-Wallis test, a non-parametric analysis of variance, was used to compare the difference in mean basal area and stem density, PPFD, and canopy openness among forest types. An alpha level of 0.05 was used as the criteria for statistical significance. SPSS ver. 12.0 (SPSS Inc. 2003) and PC-ORD ver. 4.0 (McCune and Mefford 1999) were applied to univariate, means of abiotic and biotic variables such as cover of litter and CWD, BHA and stem density, and multivariate analyses, compositional analysis, respectively.
Mean canopy openness (
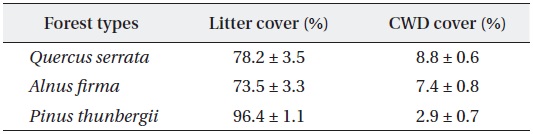
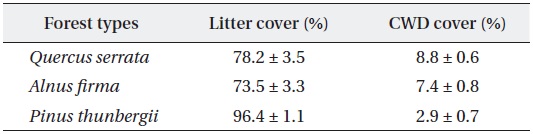
Mean and standard errors (± SD) for litter and CWD cover of studied forest types at forest floor
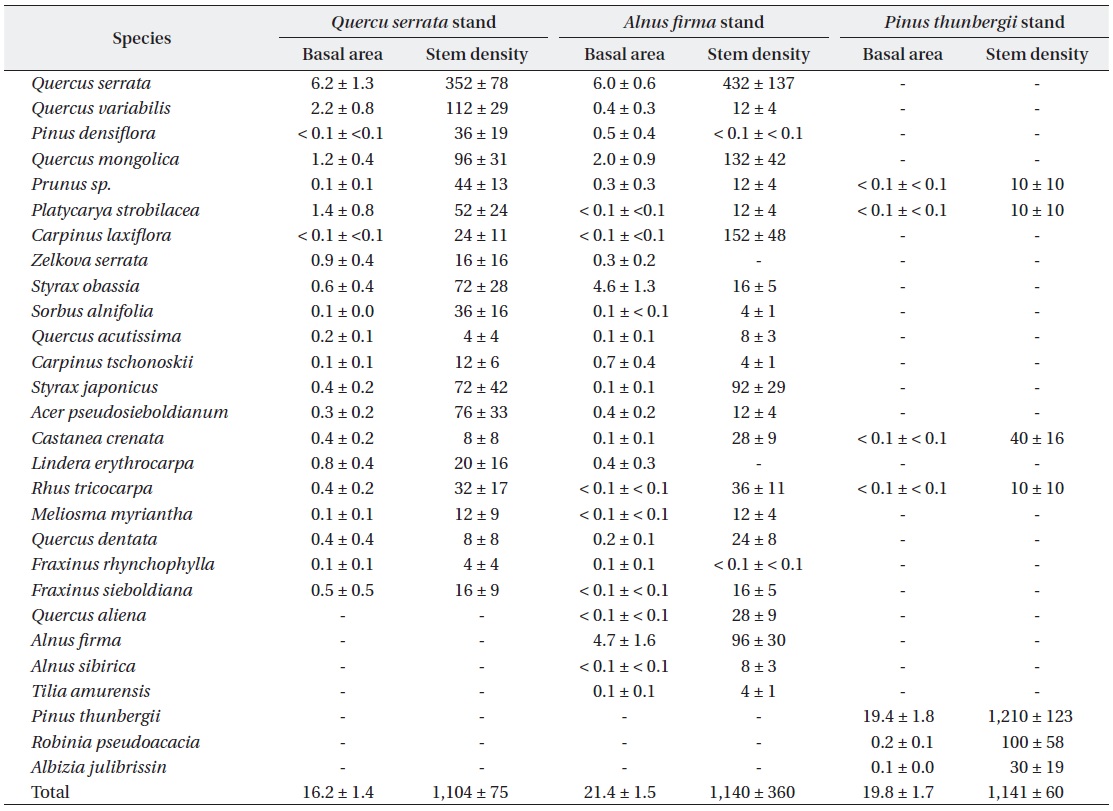
There was no significant difference on mean stem density (stems / ha,
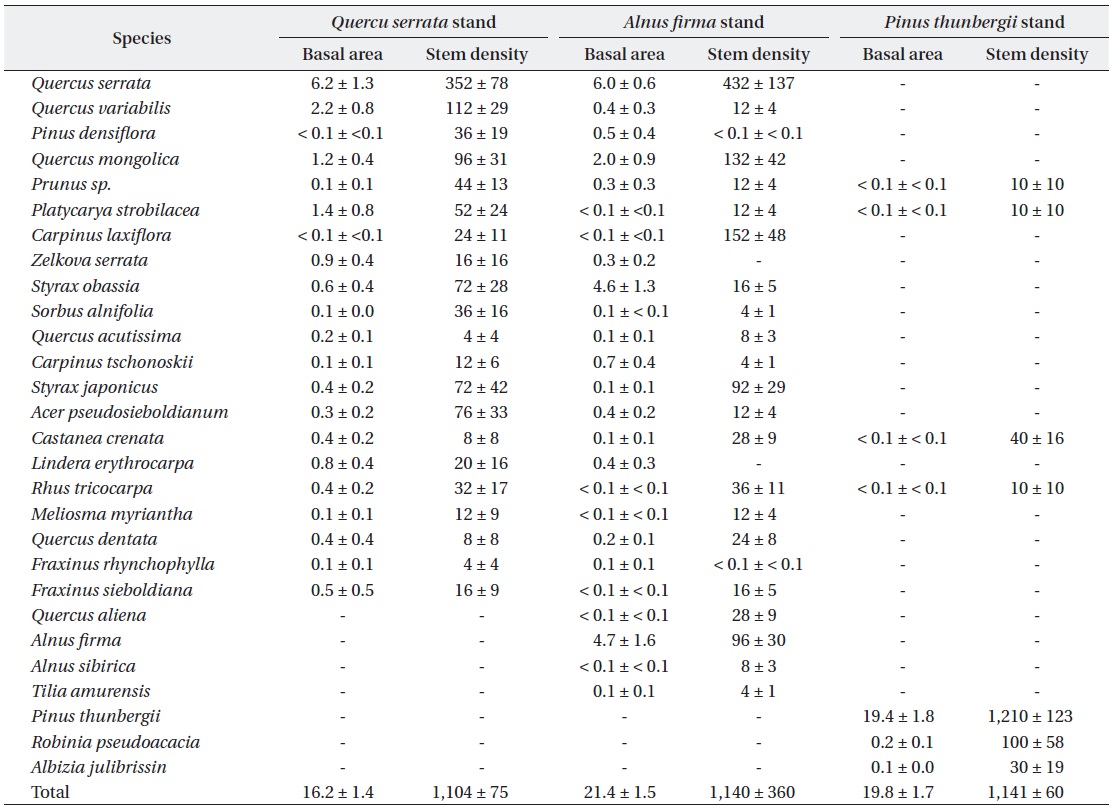
Mean basal area (m2 ha-1) and stem density (stems ha-1) for woody species greater than 5cm diameter at ground height (0.3 m) among studied stands. All values are presented with mean value and ± SE, and SE means standard error for mean. Differences in mean values of basal area (H = 5.7, p = 0.058) and stem density (H = 3.6, p = 0.162) by Kruskal-Wallis tests were not significant at 0.05 level
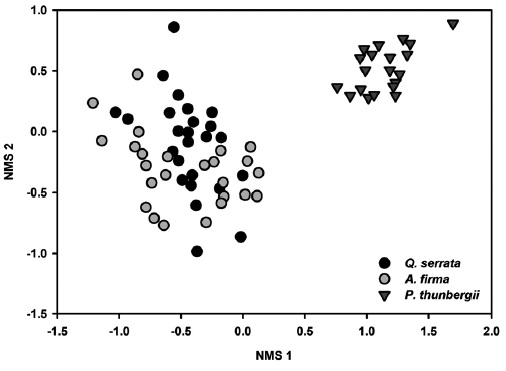
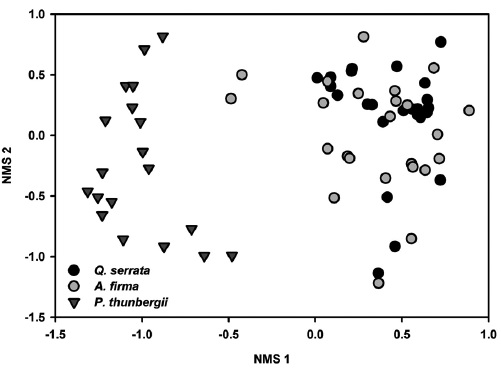
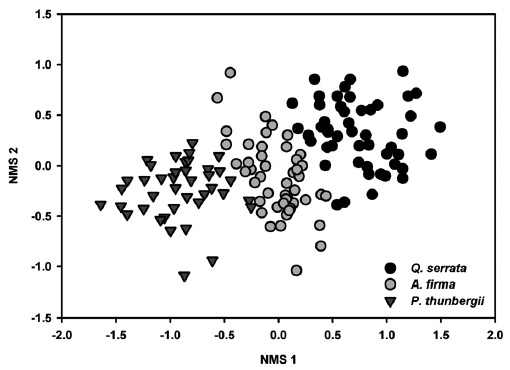
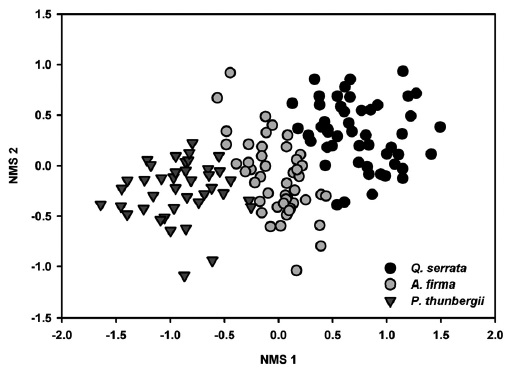
All tested MRPP exhibited significant but low differences among and between groups (Table 3). In tests among three forest types, the values of size difference were in the order of tree species composition (
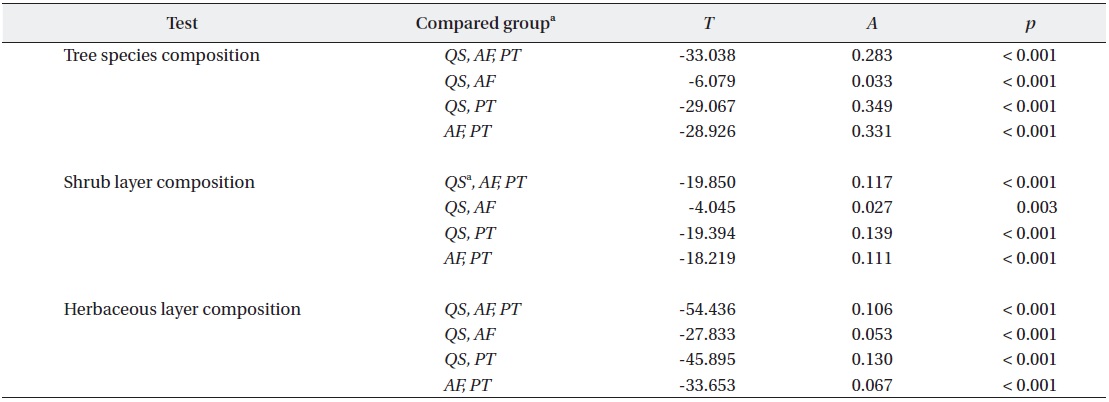
Result of the multiple response permutation procedures (MRPP) testing the differences in species composition among stand types. Samples of each group for MRPP were selected only in unmanaged sites to eliminate management noise but not for Pinus thunbergii
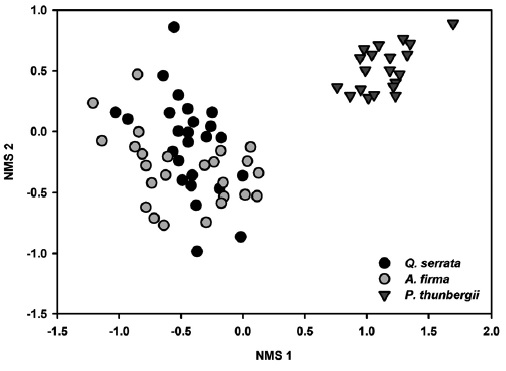
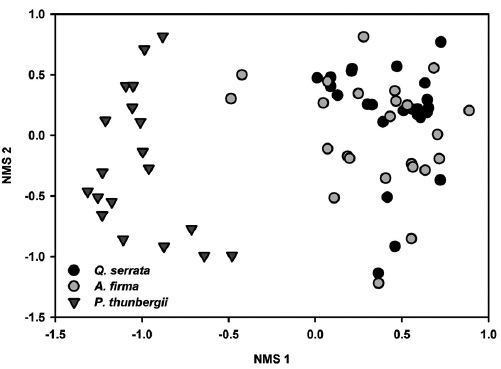
Results of NMS ordinations based on the importance values of tree species and percent cover of shrub and herbaceous layer vegetation well supported the results of MRPP (Figs. 3, 4, and 5). The compositions of tree species and shrub layer of
In ground layer, indicator species analysis (ISPAN) yielded 32 taxa showing positive associations with specific stand types (Table 4). Compared to
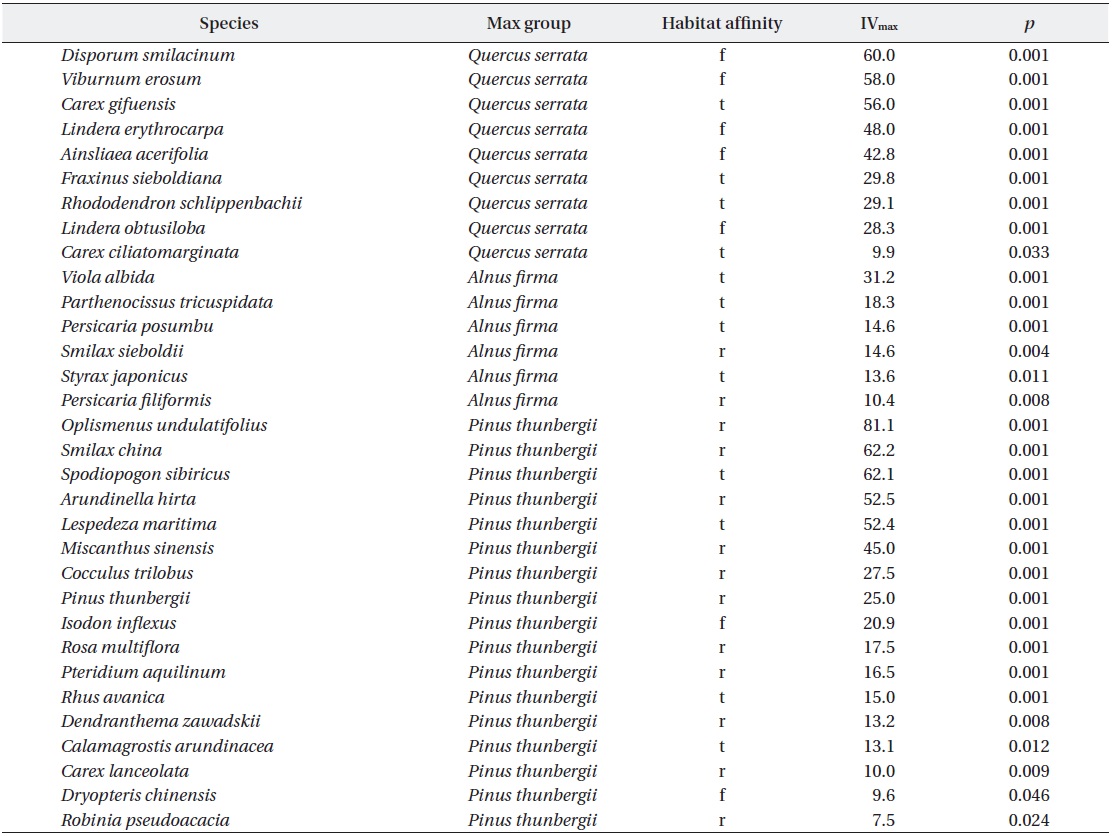
Understory species exhibiting significant associations (p < 0.05) with a particular group of stand types, based on indicator species analysis (ISPAN). Codes for habitat affinity are forest (f), transitional (t) and ruderal (r)
Overally, total of 171 species with 67 families, 121 genera, 129 species, 2 subspecies, 35 varieties and 5 forms were occurred in studied 24 sampling bands (10 m x 50 m). For each stand types, total of 110 species with 51 families, 80 genera, 84 species, 1 subspecies, 22 varieties and 3 forms were occurred in
Natural vegetation is the results of many contingent and stochastic factors (del Moral et al. 2007). During the assembly of vegetation, conditions that filters, moisture, nutrients, light and biotic pressures, immigrants change, leading to a different set of new colonist (Fattorini and Halle 2004). Therefore, a complete replica of the pre-disturbance ecosystem is not realistic and limited because much of the damages in our environment are irreversible (Choi 2004, Choi et al. 2008).
>
Local environmental effects on forest restoration
There were little differences in environmental variables and community structures between broadleaved forests of
Relatively higher differences of composition including floristic composition were detected between secondary oak and planted black pine forests. Propagule availability from landscape context such as connectedness to natural stands (Graae et al. 2003), and continuous management practices, thinning and understory removal (Graae and Heskjær 1997) in Korean black pine stands are available explanations for gaps between secondary and restored Korean alder plantations. Lee et al. (2005) also noted that native species establishment could be poor on sites isolated from natural seed sources in reforested pre-degraded forest habitat.
Stem release activities applied to black pine stands exhibited brighter environment in forest floor. Higher ground litter cover is related to higher litter production and lower decomposition rate of pine species (Mun and Joo 1994), but the effect of structural factors, stem density and stand age, was unclear because of absence of reference data on natural black pine stands. Dense and thick pine litter coverage in planted
>
Structural and compositional restoration with tree plantation
Plantation produce positive effects in landscape level and also revealed significant restorative consequences on structure and composition in pre-degraded forest land (Lugo 1992). Successive reforestation of fast-growing and N-fixing exotic
Restoration of vegetation structure was more effective in fast-growing and N-fixing
Introduction of species to degraded land is to restore ecosystem structure and function rather than to establish of species own. Compared with fully restored structural aspect of mean basal area and stem density of rehabilitated