This study investigated the occurrences and succession patterns of harmful cyanobacteria, as well as environmental factors, during a 3-year period (September 2012 to August 2015) in the upper region of the Nakdong River around Sangju weir. A total of 27 cyanobacterial taxa were observed in this study, and classified into 26 species and 1 variety belonging to 11 genera, 5 families, and 3 orders. Cell density ranged from 24 to 42,001 cells/ml, with a geometric mean of 33 cells/ml, during the survey period. The dominant orders differed depending on the survey year; order Oscillatoriales in 2013, Chroococcales in 2014 and Nostocales in 2015. An
낙동강은 총 길이 525 km, 유역면적 23,859 km2로서 국내에서 가장 길고, 유역면적이 전 국토의 24.1%를 차지하는 넓은 강이다. 낙동강 본류에는 2012년까지 진행된 낙동강 살리기 사업으로 총 8개의 보가 건설되었으며, 이 중 상주보는 낙동강 본류에 설치된 다기능 보 중 가장 상류에 위치하고 있다. 상주보가 위치한 낙동강 상류 수역은 낙동강에 위치한 보 구간 수역 중에 비교적 수질이 양호하고, 수환경이 청정한 것으로 알려져 있다(NRERC, 2014). 낙동강에서 식물플랑크톤의 종조성 및 군집구조, 녹조현상에 대한 연구는 주로 중 하류를 중심으로 이루어져왔으며(Park et al., 2015; Son, 2013; Yu et al., 2014), 상류수역에서의 식물플랑크톤 특히 녹조현상 유발 남조류를 중심으로 한 연구는 매우 미비한 실정이다(Hur et al., 2013).
녹조현상을 유발하는 남조류의 발생과 천이, 감소의 원리에 대한 이해는 남조류에 의한 녹조현상을 관리하는 측면에서 필수적이라 할 수 있다(Ma et al., 2015). 본 연구에서는 상주보 상류의 낙동강 본류 수역과 주요 유입하천을 대상으로 녹조현상 유발 남조류의 발생과 천이양상을 알아보고, 특히
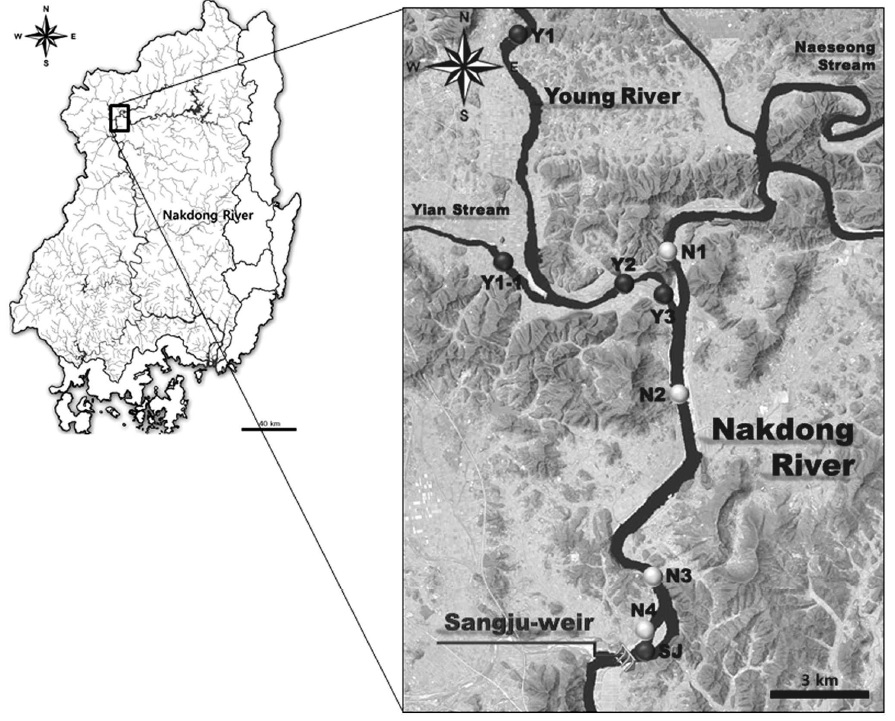
본 연구는 낙동강 상류 수계에 위치한 상주보 상류 수역을 중심으로 일반조사와 집중조사를 구분하여 실시하였다. 일반조사는 상주보 상류 500m 지점(SJ)을 대상으로 2012년 9월부터 2015년 8월까지 매주 1회 조사를 실시하였으며, 집중조사는
현장에서 채수된 식물플랑크톤 시료는 Lugol 용액으로 고정하였으며, 실험실로 시료를 옮겨 현미경(Zeiss, Axio Scope. A1)으로 검경하여 출현 조류종을 동정하였다. 정량분석은 Sedwick-Rafter chamber를 사용하여 출현 세포수를 ml당 세포수로 계수하였다(MOE, 2011). 식물플랑크톤의 동정은 한국의 조류(담수산 남조류 I, II) (NIBR, 2012a; 2012b)와 Komárek and Anagnostidis(1999, 2005)를 참고하였다. 이화학적 환경요인 중 수온, 전기전도도(EC), pH, 용존산소농도(DO)는 현장수질측정기(YSI556MPS)를 이용하여 현장에서 측정하였으며, 채수한 시료는 실험실로 운반하여 수질오염 공정시험방법(MOE, 2011)에 따라 SS, TN, TP, Chl-a를 분석하였다. 조사지점의 기상과 수리·수문에 대한 자료는 기상청(KMA, 2015) 및 한국수자원공사(Kwater, 2015)에서 제공하는 자료를 각각 참고하여 활용하였으며, 주간 누적 및 평균값을 산출하여 적용하였다. 영강 수계의 과거년도(2013년, 2014년) 수질 자료는 환경부 물환경정보시스템(MOE, 2015)를 이용하여 분석하였다.
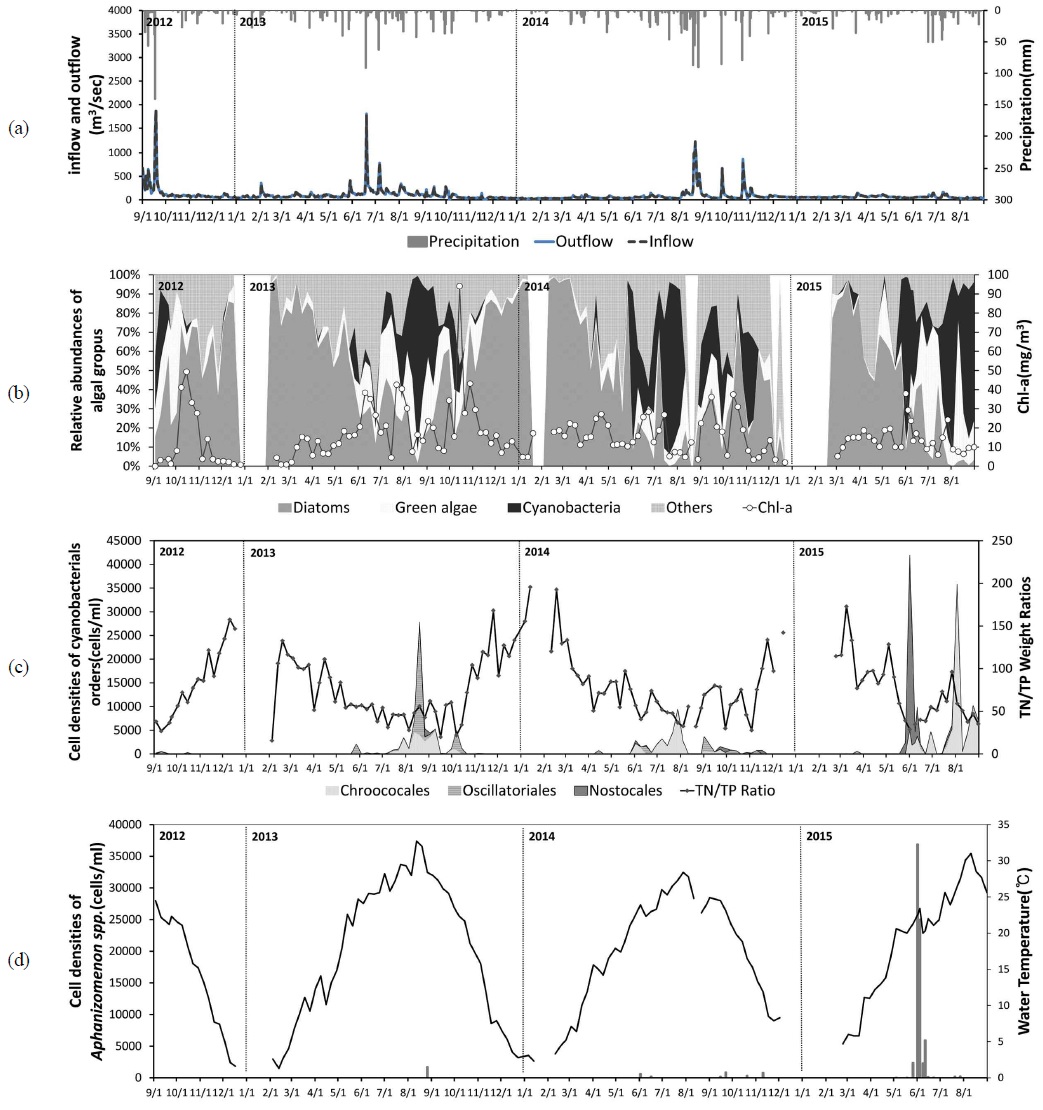
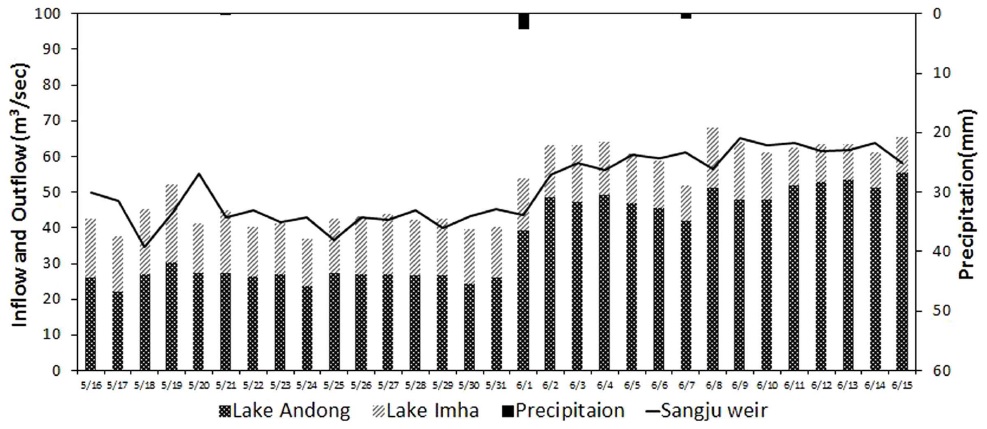
상주보가 위치한 상주 지역의 조사 기간 연평균 총강수량은 1,033 mm로 연간 강수량의 60% 이상이 하절기(6~9월)에 집중되어 나타나는 몬순 강우의 특징을 가지고 있었으며, 이로 인하여 상주보의 유입량 및 방류량 또한 하절기에 집중되고 동절기에 감소하는 변화패턴을 보이는 것으로 나타났다. 그러나 집중 강우의 시기와 빈도에 의한 변화는 연도별로 차이를 보였는데, 최대 유입량이 1,000 m3/sec를 초과하는 시기가 2013년에는 이른 장마로 인하여 초여름(6월)에 시작된 반면, 2012년과 2014년에는 늦여름(8월)에 시작되는 특성을 보였다. 한편 2015년은 강수량 부족으로 인하여 8월까지 상주 지역의 누적강수량이 537.5 mm (2013년 685.3 mm, 2014년 822.5 mm)에 불과하였으며, 상주보의 월평균 최대 유입량이 79.9 m3/sec를 초과하지 못하였다(Fig. 2(a)).
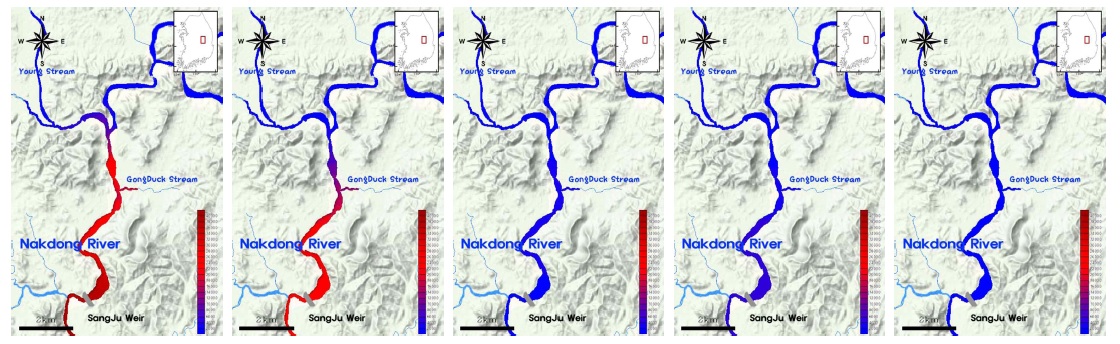
조사기간 동안 SJ 지점에서 조류 분류군별 분포는 봄과 겨울에 규조류가 우점하고, 이른 여름을 중심으로 녹조류와 기타조류가 출현하며, 늦여름에 남조류가 우점하는 전형적인 온대 수계 천이패턴을 보였으나, 남조류 발생 및 천이패턴에는 연도별로 차이를 보였다(Fig. 2(b)). 이른 장마가 시작된 2013년에는 늦은 여름(8월 이후)부터 남조류의 우점이 확인되었으나, 2014년과 2015년에는 초여름(6월)부터 남조류의 우점이 지속적으로 관찰되는 특성을 보였다. 한편, 유사지점에서 조사된 선행연구 결과를 보면 2000년의 조류 분류군별 분포는 연중 규조류(
본 연구에서 출현이 확인된 분류군 중 남조류는 총 3목 5과 11속 26종 1변종으로 분류되었으며(Table 1), 총 조사기간 동안 출현 개체밀도는 24~42,001 cells/ml (기하평균 22 cells/ml)의 범위로 확인되었다. 분류군별 개체밀도는 Chroo-coccales 목이 64~35,741 cells/ml (기하평균 9 cells/ml), Oscillatoriales 목이 52~23,464 cells/ml (기하평균 7 cells/ml), Nostocales 목이 24~36,873 cells/ml (기하평균 3 cells/ml)로 각각 확인되어,
[Table 1.] List of cyanobacterial species found at sampling stations from Sep. 2012 to Aug. 2015

List of cyanobacterial species found at sampling stations from Sep. 2012 to Aug. 2015
조사기간 동안 SJ지점에서 Nostocales 목에 의한 우점은 2015년 6월에 처음으로 확인되었으며, 이 시기의 우점종은
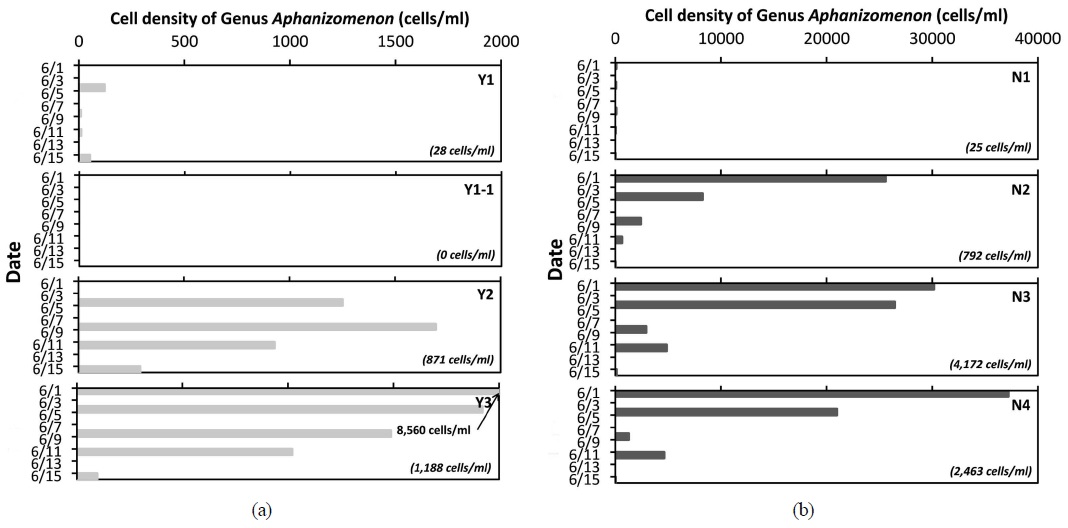
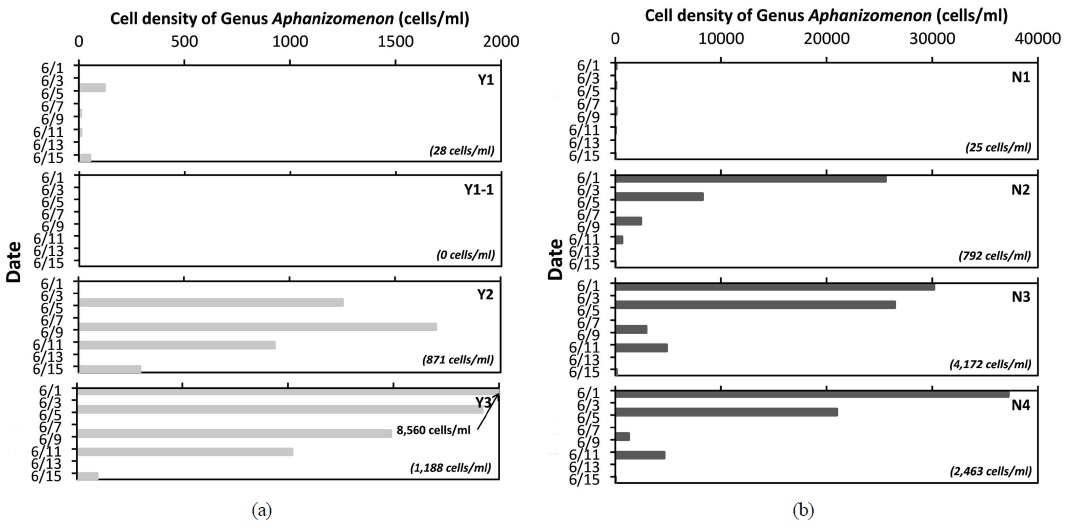
3.2. 집중조사 기간 Aphanizomenon 속의 증식 및 감소 추이
낙동강 상류 수역에서
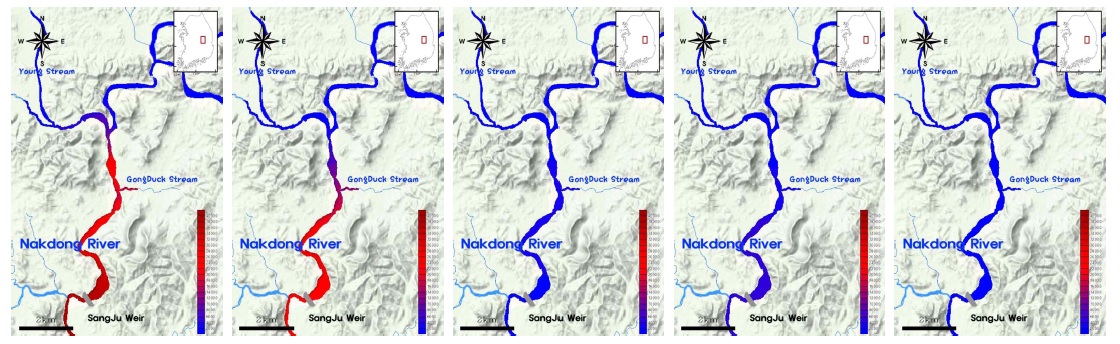
3.3. 영강에서 Aphanizomenon 속의 발생원인 분석
본 연구를 통해, 낙동강 상류 수역에서 2015년 6월에
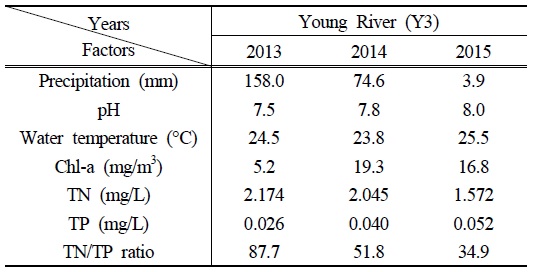
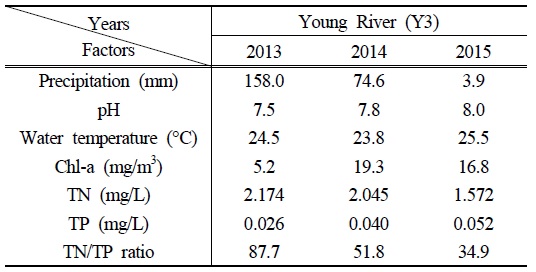
영강의 조사기간 평균 수온과 pH는 2015년에 25.5℃와 pH 8.0으로 각각 나타나, 2013년(24.5℃, pH 7.5)과 2014년(23.8℃, pH 7.8)에 비해 높았던 것으로 확인되었다. 하절기의 빠른 수온 증가와 pH의 변화는 저층에 침전되어 있는
Precipitation and physicochemical parameters at Y3 station during the Aphanizomenon bloom event (third week of May to second week of June 2015). The precipitation and physicochemical parameter values are total amounts and averages for 4 weeks, respectively
결과적으로 2015년의 영강에서는 안정적인 수리·수문학적 환경과 akinete의 발아율을 높일 수 있는 pH, 성장에 유리한 수온과 TN/TP ratio 등의 환경조건으로
상주보가 위치한 낙동강 상류 수역에서 2012년 9월부터 2015년 8월까지 3년간 남조류의 출현양상과 수환경 요인을 조사하고,