Pacific salmon are widely distributed in Far Eastern waters, where they are an important protein resource for the human population. Local consumers greatly appreciate commercially prepared salmon meat that is available in fresh and in frozen forms. The annual catch of salmon in the Far East is 400,000–500,000 ton.
By-products of salmon, including viscera, skin, head, liver, bones and roes, that are produced by seafood processing plants amount to >20% of total raw fish weight. They are usually processed into low-value products, such as animal feed, fish meal and fertilizer; some are discarded and cause water pollution.
To meet an increasing demand for better utilization of limited natural resources and reduction of environmental pollution by fish processing, modern technologies have been developed to produce a broad spectrum of food ingredients and industrial products. Enzyme technology is now available for protein recovery from waste products in the processing of salmon or underutilized species. The protein concentrates produced in this manner have generated considerable industrial interest. Hydrolysis may be used to improve or modify the physicochemical, functional, and/or sensory properties of native proteins without losing nutritional value (Kristinsson and Rasco, 2000). Protein hydrolysates from waste fish products have been produced worldwide (Chalamaiah et al., 2012). These include Pacific whiting solid wastes (Benjakul and Morrissey, 1997), and Atlantic salmon (Kristinsson and Rasco, 2000) and tuna livers (Ahn et al., 2010). Je et al. (2009) used enzymatic hydrolysis functions of commercial proteases, such as Flavourzyme, Alcalase, Protamex, and Neutrase, to produce protein hydrolysates from skipjack tuna liver, a fish by-product that is used to produce fish meal and animal feed, or discarded as processing waste.
Hydrolysis technology could also be used for better utilization of salmon livers. Our starting premise was that by developing an appropriate technology we would be able to produce a functional and nutritional ingredient giving added value to the hydrolysates of salmon. Thus, we investigated the optimum conditions, nutritional value and functionalities of enzymatic hydrolysates using salmon
The livers of frozen Pacific salmon (
[Table 1.] Characteristics of hydrolysis enzyme

Characteristics of hydrolysis enzyme
The livers were homogenized for 3 min in cold (5℃) distilled water (1:1 ratio; w/v) using a homogenizer (Ika 25T basic, IKA-Works, Wilmington, NC, USA) operating at 8000 rpm. The protein extract was adjusted to pH 7.0 using 2 N NaOH, and centrifuged at 5000 rpm for 20 min (H-3FR centrifuge, Kokusan Denki, Shizuoka, Japan) to separate the lipid layer from residues. The middle layer of centrifugate containing proteins was separated and adjusted to pH 4.0 using 0.1 N HCI, and then held in a cold room (4℃) for 30 min. The suspension was centrifuged at 7,000 rpm for 20 min. The protein precipitate was dissolved in distilled water and adjusted to pH 7.0 using 2 N NaOH. The suspension was centrifuged at 7,000 rpm for 20 min. We used the protein isolate suspension (pH 7.0) as a substrate for the proteases. The protein concentration in the protein isolate suspension was 638 mg/mL.
Solubility was determined in a protein isolate suspension dispersed in a solution of 0.1 M HCl-KCl (pH 2.0), 0.1 M citrate-phosphate (pH 3.0–5.0), 0.1 M sodium phosphate (pH 6.0–8.0) and 0.1 M glycine-NaOH (pH 9.0–11.0). The dispersions were centrifuged at 3,000 rpm for 15 min. Aliquots of filtrate were taken for determination of protein concentrations by the Biuret method (Klompong et al., 2007).
We added five volumes of distilled water to each volume of sample. The samples were then homogenized, adjusted to pH 6.0, and hydrolyzed for 1–2 h at 40℃ with 1% Protamex (w/w basis). We subsequently inactivated the enzymes in each hydrolysate (those subjected to one-step processing) by boiling at 100℃ for 10 min, after which they were held at low temperature. The inactivated hydrolysates were hydrolyzed for 1–2 h at 50℃ with 1% Alcalase (w/w basis) following adjustment to pH 8.3, and then subjected to enzyme deactivation at 100℃ for 10 min before storage at low temperature for later use. The hydrolysates subjected to two-step processing were centrifuged (5,000 rpm, 20 min), filtered and freezed-dried for later use in anaytical solutions.
>
Determination of the degree of hydrolysis
Degree of hydrolysis (DH) was calculated on a percentage bases by the trichloroacetic acid (TCA) ratio method described by Hoyle (1994). After hydrolysis, 20 mL of protein hydrolysate were added to 20 mL of 20% TCA (w/v) to produce 10% TCA soluble material. The mixtures were left to stand for 30 min to allow precipitation, and then centrifuged at 10,000 rpm for 10 min. The supernatant was analyzed for protein content following the procedures of Lowry et al. (1951). Samples from the hydrolysate were also analyzed for protein content. The degree of hydrolysis was calculated as follows:
% DH = (soluble protein in 10% TCA / total protein in the sample) × 100
1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging activity
We measured the free-radical-scavenging activity of hydrolysate using the DPPH free radical method (Park et al., 2005). We mixed 0.1 mL sample volumes of varying concentration with 0.15 mL of DPPH (0.4 mM) solution in ethanol. Absorbance at 517 nm was measured using a microplate reader (Powerwave XS, BioTek, Winooski, VT, USA; UV 1800 spectrophotometer, Shimadzu, Kyoto, Japan). Scavenging activities were expressed as the IC50, which was the concentration of the test sample that inhibited 50% of substrate activity. Vitamin C was used for comparison with DPPH radical scavenging activities of the samples.
Antifibrinolytic activity was determined following the method of Masahiro (2004); briefly, 80 μL of 1.75 μM fibrinogen were mixed with 20 μL of 7.5 U/mL thrombin (from normal human plasma [T6884-100UN]) and 20 μL of sample; the combination was then injected into 96-well plates. Subsequently, we recorded the time to clotting, and then measured the absorbance of the mixture (at 655 nm) at 37℃ over a 3 h period (at 10 min intervals). One unit was defined as the enzyme level required to decrease absorbance by 0.1 per 60-min reaction time.
>
α-glucosidase inhibitory activity
Reaction mixtures, each containing 1-μL α-glucosidase (2 units m/L, 0.1 mol/L, potassium phosphate buffer, pH 6.8) pre-mixed with 1 μL of sample solution (0.01–200 μg/L in 10% dimethyl sulfoxide [DMSO]) in 95 μL of 0.1 mol/L potassium phosphate buffer (pH 6.8), were injected into 96-well plates. Following incubation for 20 min at 37.5℃, we added 3 μL of p-nitrophenyl-β-D-glucopyranoside (PNPG) solution (3 mmol/L) as substrate to the mixture to initiate the reaction. The reaction mixture was incubated at 37.5℃ for 20 min, after which we added 100 μL of 1 mol/L Na2CO3 solution to terminate the reaction. The amount of released
>
α-amylase inhibitory activity
Twenty microliters of α-amylase solution and 50 μL of sample solution were mixed and incubated for 5 min at 37℃ with 430 μL of 0.5% starch solution in phosphate-buffered saline (PBS). The reactions were allowed to proceed for 15 min at 37℃ following addition of 500 μL of 3,5-dinitrosalicylic acid (DNS) reagent; subsequently, we held reaction vessels in a water bath at boiling point for 5 min, and then cooled them to room temperature for determination of absorbance at 540 nm. The α-amylase solution comprised α-amylase (35 U/mg) in 5 mg/mL in distilled water. The DNS reagent was made up as follows: 1 g of 3,5-dinitrosalicylic acid was dissolved in 60 mL distilled water, to which we added 1 g of NaOH; the mixture was subsequently heated in a water bath at 45℃; 12-g Na-K tartrate, 0.2-g phenol and 0.05-g sodium sulfite were then added; after the compounds had dissolved, the volume was made up to 100 mL and filtered. The prepared reagent was storable in darkness for 7 days.
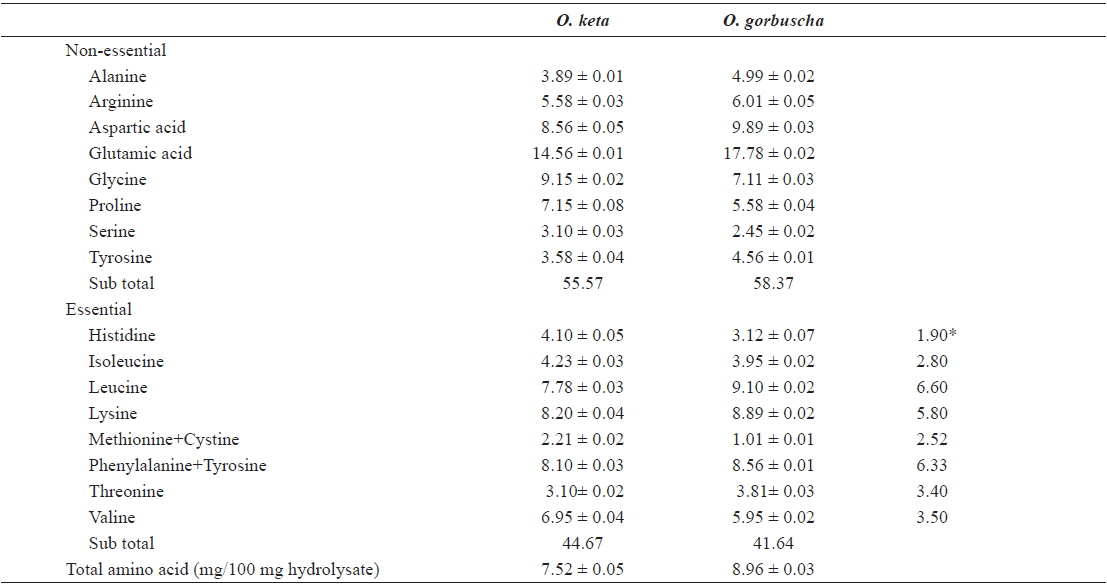
The nutritional value of any ingredient depends on the ability of its proteins to fulfill the needs of consumer organisms. Essential amino acid compositions and chemical scores (computed in relation to the FAO/WHO reference protein) were used to evaluate the nutritive value of proteins. We determined the amino acid compositions of salmon liver hydrolysates by digestion in 6 N HCl/0.1% phenol solution at 110℃ for 20 h following the Pico–Tag method of White (1986). Amino acid compositions of salmon liver hydrolysates prepared with Alcalase enzyme are listed in Table 2. Amino acid profiles of liver hydrolysate were generally similar between
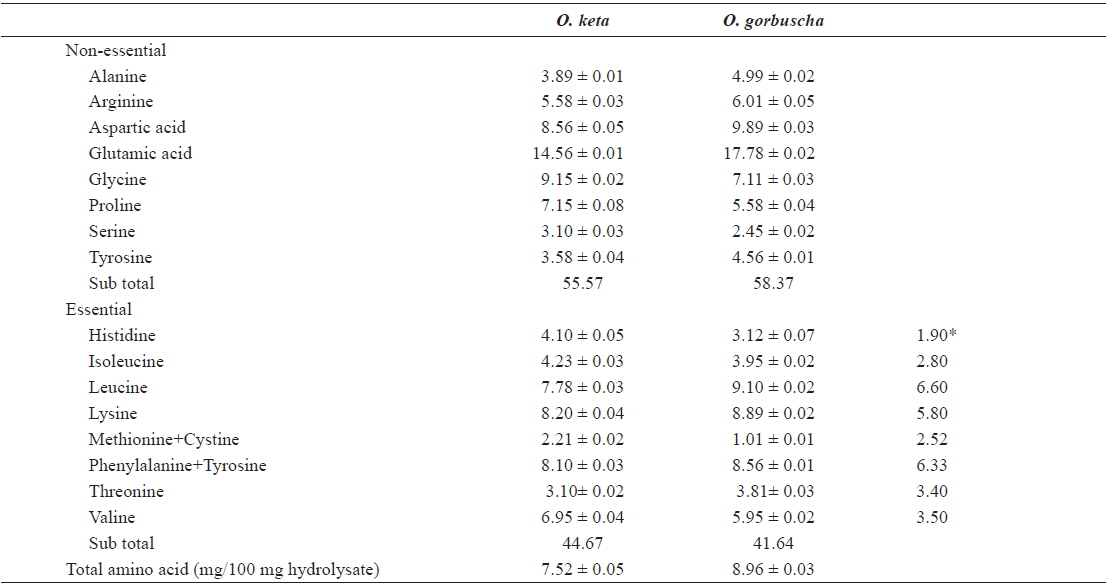
Amino acid composition (% w/w) of the active fractions of salmon liver hydrolysates from Oncorhynchus keta and Oncorhynchus gorbuscha
Differences in amino acid composition between the hydrolysates may be attributed to differences in the specificity of proteases. Chemical scores indicated that all amino acids (other than methionine and threonine) were present in adequate or excess quantities. An elevated ratio of hydrophobic amino acids (arginine, tyrosine, isoleucine, methionine, phenylalanine, and valine) reportedly improves antioxidant activities (Chen et al., 1996), and cytotoxic and anticancer effects (Hoskin and Ramamoorthy, 2008). The hydrophobic amino acids proportions in hydrolysates prepared from
>
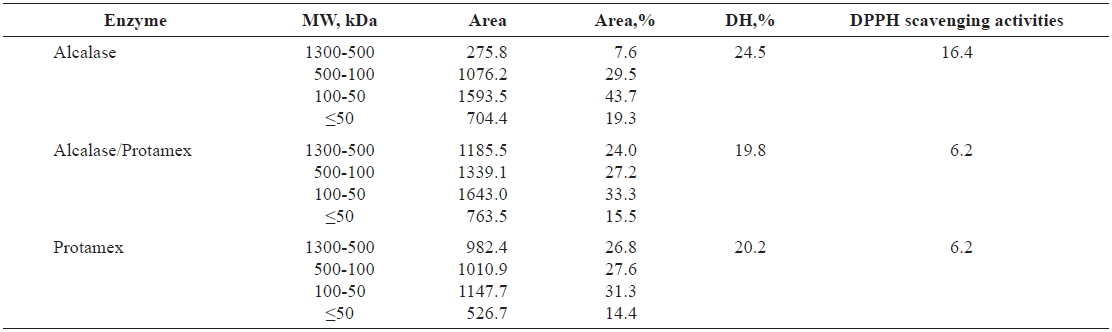
Molecular weight (MW) distribution of Pacific salmon liver hydrolysates
The molecular sizes of proteins and peptides are important physicochemical parameters that are correlated with biological activity. The MW distributions of the peptides in salmon liver hydrolysates are listed in Table 3. Alcalase was the most efficient enzyme in terms of degrading liver extract proteins; nearly all high-MW proteins were cleaved to produce low-MW peptides, as indicated by our estimates of DH.
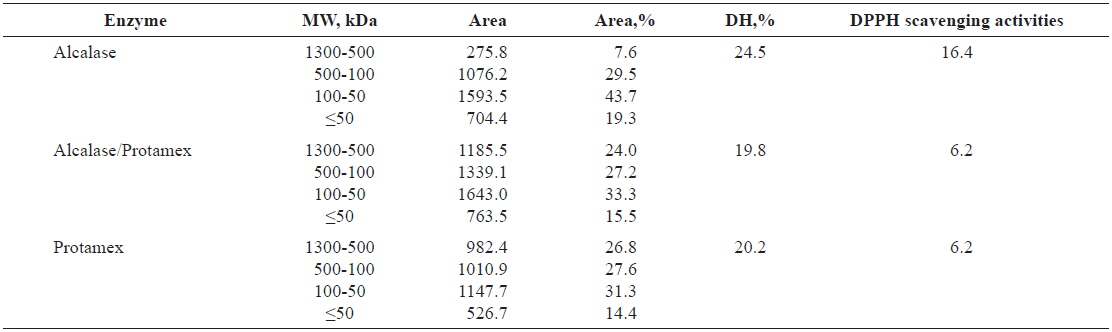
Relative percent of the peptides in HPLC in total area (%) of salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver hydrolysates
Hydrolysates in the 500–1,300-kDa MW range produced by Alcalase exhibited the highest DPPH-scavenging activities among the hydrolysates. Tuna liver protein has been treated to liberate low-MW bioactive peptides by hydrolysis with Flavourzyme, and subsequent digestion with the endopeptidases Alcalase, Neutrase and Protamex (Ahn et al., 2010). The tuna protein was fractionated into four MW categories to identify biological activities. All fractionated hydrolysates in the MW range above 10 kDa produced by the enzyme combinations Alcalase+Flavourzyme, Neutrase+Flavourzyme and Protamex+Flavourzyme had elevated reducing power (Ahn et al., 2010).
The proportion of released peptides exceeding 500 kDa was low when hydrolysates were treated with Alcalase, but the proportions in the 100–500-kDa and <50-kDa ranges were elevated. Thus, liver hydrolysate protein likely has large numbers of sites suitable for the enzymatic cleavage activity of Alcalase; the protein was less suitable for cleavage by Protamex and the Protamex/Alcalase combination. Our results were congruent with other studies on Atlantic cod viscera (Aspmo et al, 2005), sea cucumber (Forghani and Ebrahimpour, 2012) and red salmon heads (Sathivel et al, 2005), which demonstrated the efficacy of Alcalase in protein cleavage for producing small peptides.
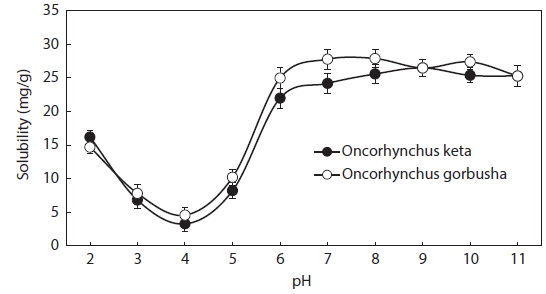
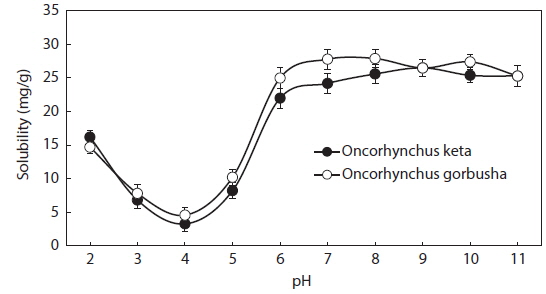
Protein solubility is an important molecular property that also influences other functional properties. The solubility profiles of salmon liver proteins in relation to pH are shown in Fig. 1. Solubility was calculated as the protein concentration in the supernatant after pH adjustment. The protein concentration in the protein isolate suspension was 638 mg/mL.
Salmon liver proteins had very low solubilities in the pH range between 3.0 and 5.0, with minimal values at pH 4.0 (isoelectric point; ca. 95% of the proteins precipitated at this pH). Solubilities increased as pH shifted in either direction away from the isoelectric point. Thus, the protein in salmon liver should be solubilized in the pH range 2.0–3.0, and precipitated at pH 4.0 for optimum recovery using acid-aided processing. The protein may also be solubilized in the pH range 6.0–9.0 when using alkali-aided processing. Minimal solubility near the isoelectric pH promotes aggregation and precipitation via hydrophobic interactions (Damodaran, 1996).
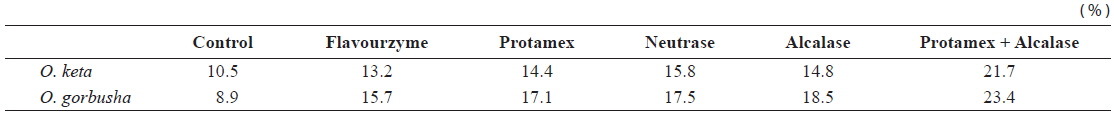
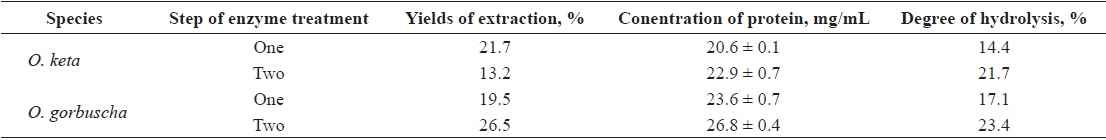
Fish protein hydrolysates have attracted much recent attention from food biotechnologists because of the availability of large quantities of raw material with high protein content, good amino acid balance and bioactive peptides (antioxidant, antihypertensive, immunomodulatory and antimicrobial peptides) (Chalamaiah et al., 2012). Degree of hydrolysis is the most widely used measure for comparing protein hydrolysates. Applications of enzyme technology for protein recovery from salmon processing have the potential to produce a broad spectrum of food ingredients and industrial products. We hydrolyzed salmon liver extract for 1 h using four commercially available enzymes. Enzyme activities were high in the pH range 5.5–8.0, highest at pH 7.0, and very low at pH 10.0.
The DHs achieved with the Protamex/Alcalase complex were 21.7 and 23.4% for
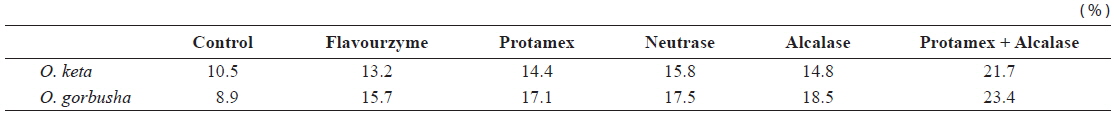
Degree of hydrolysis of various enzyme-treated hydrolysates from salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver
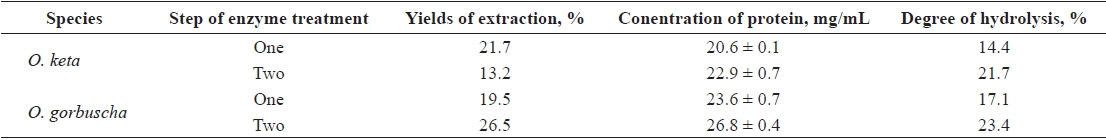
Yields and protein concentration and DH of salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver hydrolysates
Industry adoption of by-product recovery and utilization procedures will depend on a clear economic benefit analysis showing that dumping of wastes is a disadvantageous alternative (Kristinsson and Rasco, 2000). High value improvements of food protein functionalities are often achieved by enzymatic hydrolysis of proteins; partial hydrolysis is especially valuable in this regard (Kuipers et al., 2005).
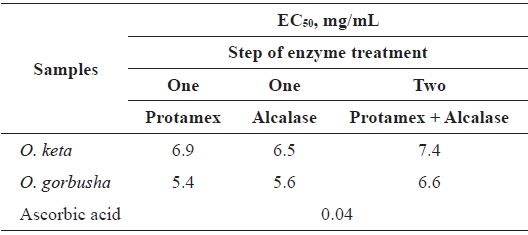
We examined DPPH-scavenging activities to determine
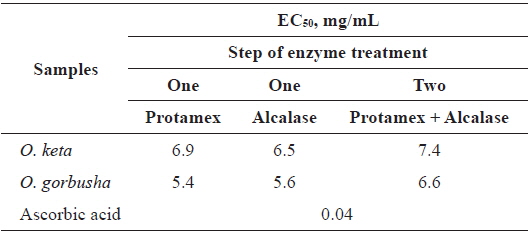
DPPH scavenging activities (EC50) of various enzyme-treated hydrolysates from salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver
The EC50 range of hydrolysate from
Tuna liver protein hydrolysates have excellent reducing power and antioxidant abilities against DPPH, hydrogen peroxide and hydroxyl radicals (Ahn et al., 2010). These antioxidant activities depended on the proteolytic enzymes used, and on the MW ranges of the fractionated hydrolysates (Ahn et al., 2010). These findings are congruent with our results showing that salmon liver hydrolysates have good DPPH scavenging activities when treated with Protamex/Alcalase in a two-step process. Thus, salmon liver is a potential source of value-added process products.
The incidence of thrombotic diseases, including atherosclerosis and stroke, is related to platelet coagulation, fibrin degradation and the regulation of the coagulation. Thrombosis is initiated by the adhesion, activation and aggregation of platelets injured by the vascular endothelial cells, and promoted by the activation of the blood coagulation system. Platelets are activated by the release of platelet ADP and thromboxane A2 (TXA2). Platelet coagulation is promoted when a platelet membrane GPIII receptor combines with fibrinogen (Yang et al., 2007; Yang et al., 2012; Fan et al., 2009).
A summary of the anti-thrombotic effects of hydrolysates extracted from salmon liver is presented in Table 7. The salmon liver hydrolysate produced by one-step enzymatic action had an antithrombotic efficacy of 0.0173 U at a papain concentration of 10 μg/mL (Table 5).

Antithrombotic effect of hydrolysate extracts from salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver
>
α-Amylase and α-glucosidase inhibitory activities
Diabetes is a metabolic disease with chronic complications, such as stroke, myocardial infraction, angina pectoris, optic nerve damage, foot ulcers. Hyperglycemic disease is longlasting and much feared by patients (Cha et al., 2010). The incidence of diabetes reaches 10% in the population of Korea, where patients with type 2 diabetes account for 90% of diabetics (Lee et al., 2006)
Inhibition of key enzymes metabolizing carbohydrate, such as α-amylase and α-glucosidase, can effectively delay the absorption of glucose, and thereby blunt the rise in postprandial plasma glucose (Tundis et al., 2007; McDougall and Stewart, 2005).
To assess the blood glucose control functions of salmon liver hydrolysate, we tested α-amylase inhibition (Table 8). The hydrolysates had inhibitory activities of 11.5% and 44.7% at 0.1 and 1.0 μg/mL, respectively. Both salmon species liver hydrolysates had lower inhibitory activity than Acarbose at 0.1 μg/mL (positive control).

α-Amylase inhibition effect of hydrolysate extracts from salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver
We also measured the α-glucosidase enzyme inhibition effect as a blood glucose control (Table 9). The hydrolysates had inhibitory activities of 7.8% and 46.5% at 0.1 and 1.0 μg/mL, respectively. Acarbose at 0.1 μg/mL was used as a positive control. The hydrolysate inhibitory activities α-of glucosidase and α-amylase were therefore similar.

α-Glucosidase inhibition effect of hydrolysate extracts from salmon Oncorhynchus keta and Oncorhynchus gorbuscha liver
Overall, we found that salmon liver hydrolysates may have useful applications in the production of functional foods. Of the proteases tested, Alcalase was the most efficient for production of hydrolysate with the highest DPPH-scavenging activity. The hydrolysates had balanced amino acid compositions and contained several low-molecular-weight peptides, which are likely bio-active. These activities are the subject of ongoing research in our laboratory.